
7.3: Oxidative Phosphorylation
- Page ID
- 16452

Oxidative phosphorylation is the mechanism that by which ATP captures the free energy in the mitochondrial proton gradient. Most of the ATP made in aerobic organisms is made by oxidative phosphorylation, rather than by substrate phosphorylation (the mechanism of ATP synthesis in glycolysis or the Krebs cycle). Some aerobic chemistry may have evolved in response to the toxic effects of rising environmental oxygen levels. Later elaboration of respiratory metabolism were undoubtedly selected because it turns out to be more efficient at making ATP than anaerobic fermentations such as ‘complete’ glycolysis. In other words, oxidative phosphorylation fueled by electron transport is more efficient that substrate-level phosphorylation. Oxidative phosphorylation couples controlled diffusion of protons through the cristal membrane ATP synthase to ATP production.
To summarize here, the movement of electrons down the electron transport chain fuels three proton pumps that establish a proton gradient across the cristal membrane that stores free energy. We say that the proton gradient has a proton-motive force, resulting from the difference in proton concentration (an H+ or pH gradient) and a difference in electric potential. The use of this force to make ATP is regulated.
Conditions in the cell control when the energy stored in this gradient will be released to make ATP. The switch that allows protons to flow across the cristal membrane to relieve the proton gradient is an ATP synthase, a tiny, complex enzymatic protein motor. For a clear discussion of this complex enzyme, see P. D. Boyer (1997) The ATP synthase – a splendid molecular machine. Ann. Rev. Biochem. 66:717-749.
The capture of free energy of protons flowing through the complex is summarized below.
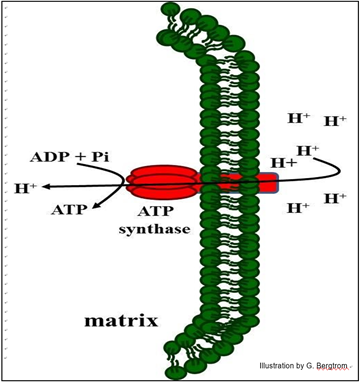
In mitochondria, the protons pumped out of the mitochondrial matrix (using the free energy released by electron transport) can then flow back into the matrix through the ATP synthase. If the three ETC sites in the cristal membrane that actively transport protons are proton pumps, then the cristal membrane ATP synthase complexes function as regulated proton gates that catalyzes ATP synthesis when protons are allowed to flow through them. For their discovery of the details of ATP synthase function, P. D. Boyer and J. E. Walker shared the Nobel Prize in Chemistry in 1997.
165 Proton Gates Capture Proton Gradient Free Energy as ATP
The ratio of ATP to ADP concentrations regulates proton flow through the ATP synthase gates is regulated. A high ATP/ADP ratio in the mitochondrial matrix indicates that the cell does not need more ATP, closing the proton gate remains closed so that the proton gradient cannot be relieved. On the other hand, a low ATP/ADP ratio in the matrix means that the cell is hydrolyzing a lot of ATP…, and that the cell needs more. Then the proton gate opens, and protons flow through cristal membrane ATP synthases back into the matrix along a concentration gradient. As they flow, they release the free energy that powers a protein motor in the enzyme that in turn, activates ATP synthesis. Recall that according to the endosymbiotic theory, aerobic bacteria are the evolutionary ancestor to mitochondria; in fact, the cell membrane of aerobic bacteria house an ETC and chemiosmotic mechanism of ATP generation much like that in mitochondria.
Proton gradients do not only power ATP synthesis, but can also power cellular work quite directly. The well-known example is the bacterial flagellum driven directly by proton flow through cell membrane proton gate/molecular motor complex (below).
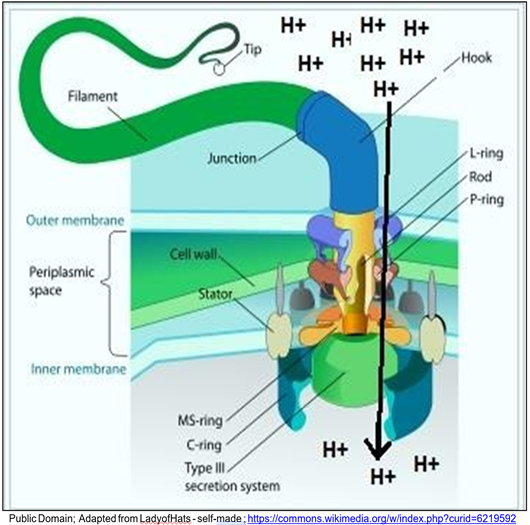
Electron transport in the cell membrane creates the gradient, and relief of the gradient directly powers the flagellum.
Just as we did for glycolysis, we can count the ATPs and see how much free energy we get from aerobic respiration, i.e., the complete oxidation of glucose. You can see this in the link below.