
5.1: Tissues
- Page ID
- 18004

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)From now on, we will frequently use multiple names of plants\(_2\) group, they are summarized on Figure \(\PageIndex{1}\), and in more details—on Figure 6.1.1.
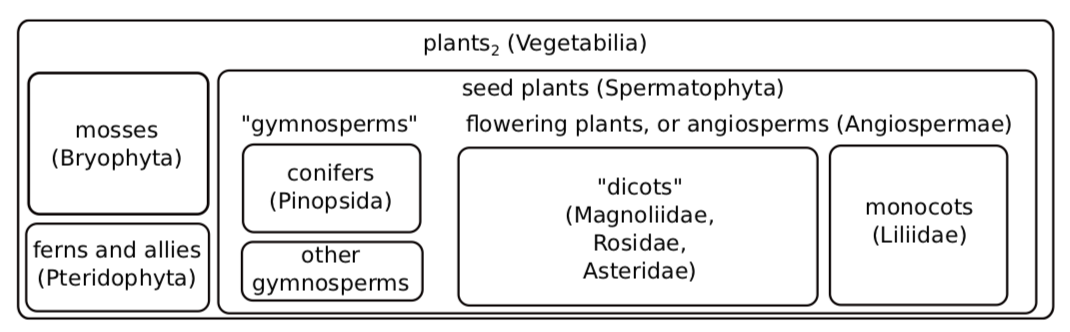
Epidermis and Parenchyma
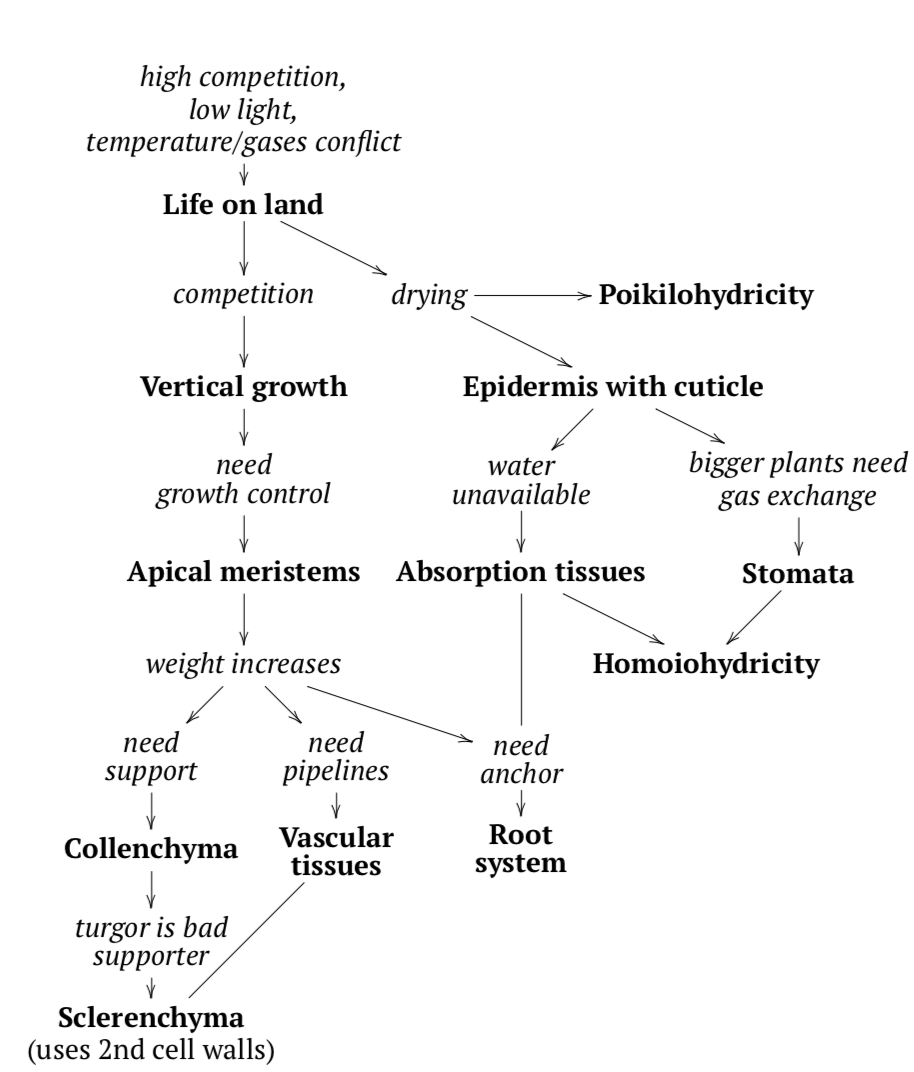
Why did plants go on land? In order to escape competition with other plants for resources like the sun and nutrients, but also to obtain much more sunlight that was otherwise seriously reduced underwater. The move to land also helped plants escape predators. Lastly, plants benefited from this change because they escaped from the temperature-gases conflict: warmer temperatures are good for organisms but significantly decrease the amount of gases diluted in water.
Although this action solved several problems, it also raised new issues that needed to be dealt with. The most important was the risk of drying out. To combat this, plants developed their first tissue: epidermis (complex surface tissue) covered with a cuticle (plastic-like isolation layer) which served a purpose similar to a plastic bag. For the really small (millimeters) plant it is enough because, in accordance to surface / volume law (i.e., when body size grows, body surface grows slower then body volume (and weight)), they have high relative surface, and diffusion can serve for gas exchange. However, bigger plants also need to exchange gases, and they developed stomata which served as a regulated pore system. The remaining cells became second tissue: parenchyma (tissue or cell type of spherical, roughly connected living cells) or ground tissue (same as parenchyma (see) but only applied for tissue), or main tissue).
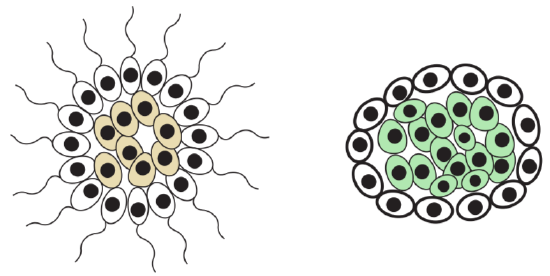
Another response (Figure \(\PageIndex{3}\)) for drying was a development of poikilohydricity (see below), the ability to hibernate in (almost) dried condition. As hibernation is generally dangerous since it requires “system restart”, that evolutionary route did not become the main. Tissue is a union of cells which have common origin, function and similar morphology. Tissues belong to organs: organ is a union of different tissues which have common function(s) and origin. Plants have simple and complex tissues. The simple tissues (tissues with uniform cells) are composed of the same type of cells; complex tissues (tissues with more than one type of cells) are composed of more than one type of cell, these are unique to plants.
Parenchyma (Figure \(\PageIndex{4}\)) are spherical, elongated cells with a thin primary cell wall. It is a main component of young plant organs. The basic functions of parenchyma are photosynthesis and storage. Parenchyma cells are widespread in plant body. They fill the leaf, frequent in stem cortex and pith and is a component of complex vascular tissues (see below). Contrary to parenchyma (which is a simple tissue), epidermis is a complex tissue composed of epidermal and stomata cells. Its main functions are transpiration, gas exchange and defense.
As it seen here, plants acquired tissues in a way radically different from animals (Figure \(\PageIndex{2}\)): while plants regulate gas and water exchange in response to terrestrial environment, animals actively hunt for food (using kinoblast tissues) and then digest it (with pagocytoblast tissue).
Supportive Tissues: Building Skyscrapers
When more and more plants began to move from the water to the land, competition once again became a problem (Figure \(\PageIndex{3}\)). To solve this, plants followed “Manhattan solution”: they grew vertically in order to be able to escape competition for the sunlight and therefore must develop supportive tissues.
Collenchyma (Figure \(\PageIndex{4}\)) is living supportive tissue that has elongated cells and a thick primary cell wall. Its main function is the mechanical support of young stems and leaves via turgor.
Sclerenchyma (Figure \(\PageIndex{4}\)) is a dead supportive tissue that consists of long fibers or short, crystal-like cells. Each cell has a thick secondary wall that is rich in lignin. Its main function is a support of older plant organs, and also hardening different parts of plants (for example, make fruit inedible before ripeness so no one will take the fruit before seeds are ready to be distributed). Without sclerenchyma, if a plant isn’t watered, the leaves will droop because the vacuoles will decrease in size which lowers the turgor. Fibers inside phloem (see below) are sometimes regarded as a separate sclerenchyma.
Three times in their evolution plants found the new application for lignin or similar polymers: at first, similar chemicals covered the spore wall which was an adaptation to the spore distribution with wind. Then similar chemicals were used to make cuticle, “epidermal plastic bag” to prevent transpiration outside of stomata. Finally, with acquiring of sclerenchyma, plants found how to use dead cells with completely lignified cell walls.
By the way, stomata likely had a similar fate, they historically appeared on sporangia to help them dry faster and release spores effectively. Regulation of transpiration is their second function.
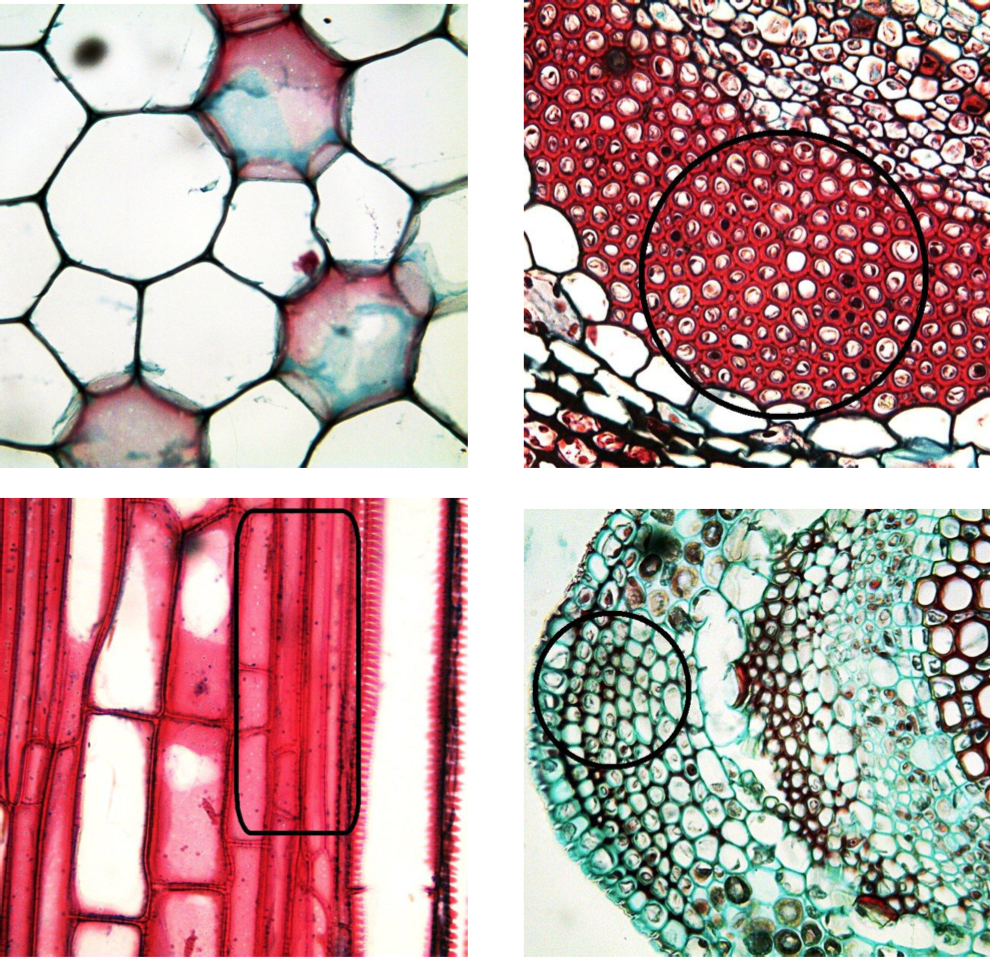
Cell types and tissues
“Parenchyma” and “sclerenchyma” terms are freShoot systemquently used in two ways: first, to name tissues (or even classes of tissues) which occur in multiple places of the plant body, and second, to name the cell types which are components of tissues. Therefore, it is possible to say “parenchyma of stem”, “parenchyma of stem pith”, “parenchyma of xylem” and even “leaf mesophyll is a parenchyma”.
Meristems: the Construction Sites
Plant growth requires centers of development which are meristemssites of cell division. Apical meristems are centers of plant development located on the very ends of roots (RAMroot apical meristem) and stems (SAMstem apical meristem). They produce intermediate meristems (like procambiumintermediate meristem developing into cortex, pith and procambium) which form all primary tissuestissues originated from RAM or SAM (optionally through intermediate meristems). The lateral meristemcambium, meristem appearing sideways or cambium originates from the procambium which in turn originates from apical meristems. It usually arises between two vascular tissues and its main functions are thickening and producing secondary vascular tissuessecondary phloem and secondary xylem (Figure \(\PageIndex{5}\)).
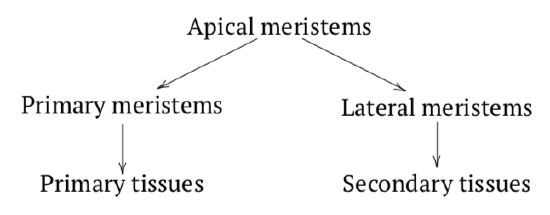
Other meristems include: intercalary which elongate stems from the “middle”, marginal which are located on margins which are responsible for leaf development and repair meristems arising around wounds, they also control vegetative reproduction.
Vascular Tissues
Bigger plants escaped from competition and performed effective metabolism. However, with all the growth the plants went through, their size became too big for slow symplastic plasmodesmata connections. Another, filter paper-like apoplastic transport was also not powerful enough. The solution was to develop vascular tissues, xylem and phloem (Figure \(\PageIndex{6}\), Figure 5.5.1).
The main functions of xylem are the transportation of water and mechanical support. The xylem tissue transporting water may be found either in a vascular bundle or a vascular cylinder. The three types of xylem cells are tracheary elements (these include tracheids
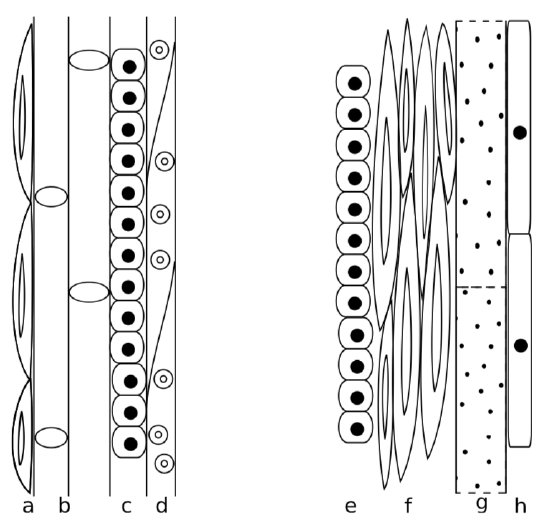
and vessel member), fibers, and parenchyma. Xylem elements, except for the parenchyma, are rich in lignin and are main components of wood. Tracheids are closed on both ends and connected with pits whereas vessel members are more or less open and connects via perforations. Tracheids, vessel members and fibers are dead cells. Xylem parenchyma, on the other hand, is alive.
Pits of tracheids consist of a pit membrane and the torus in a center, there are no openings. The presence of tracheids and/or vessel elements has evolutionary significance. Vessels (made of vessel members) are more effective; consequently, more “primitive” plants have more tracheids whereas more “advanced” have more vessel members. As an example, gymnosperms have only tracheids while most flowering plants have tracheids and vessel members. Individual development also mimics this evolutionary trend. Younger flowering plants have more tracheids whereas mature plants have more vessel members. Primary xylem mostly has tracheids and vessels with scalariform perforations whereas secondary xylem (which originates from cambium) consists mostly of vessels with open perforations. The common name for secondary xylem is wood.
It is a mistake to think that tracheids are better than vessels. In fact, the main problem is frequently not too slow but too fast water transport. Tracheids have an advanced connection system (called torus) which has the ability to close pore if the water pressure is too high and therefore more controllable. Leaking would be less dangerous in tracheids. And in water-poor environments (like taiga in winter), plants with tracheids will have the advantage. Contrary, having vessels is like to have race car for ordinary life; only flowering plants “learned” how to use them effectively.
Dead cells are useful but hard to control. However, if xylem transport needs to be decreased, there is a way. Xylem parenchyma cells will make tyloses“stoppers” for tracheary elements made by parenchyma cells (“stoppers”) which will grow into dead tracheary elements and stop water if needed. Many broadleaved trees use tyloses to lower xylem transport before the winter.
The phloem tissue transporting sugars generally occurs adjacent, or right next to, the xylem, with the xylem facing the inner part of the plant and the phloem facing the outer part of the plant. The main functions of the phloem are the transportation of sugars and mechanical support. The four types of phloem cells are: sieve tube cells , companion cells, fibers (the only dead cells in phloem), and parenchyma. Sieve tube cells of flowering plants have cytoplasm flowing through perforations (sieve plates) between cells but do not contain nuclei. Companion cells will make proteins for them. However, in gymnosperms and more “primitive” plants there are no companion cells at all, so sieve tube cells do contain nuclei. This is comparable to red blood cells in vertebrates: while mammals have them anucleate, erythrocytes of other vertebrates contain nucleus. The secondary phloem generally has more fibers than the primary phloem.
This small table summarizes differences between xylem and phloem:
| Xylem | Phloem | |
| Contains mostly | Dead cells | Living cells |
| Transports | Water | Sugar |
| Direction | Up | Down |
| Biomass | Big | Small |
Periderm
*Peridermperidermsecondary dermal tissue is a secondary dermal tissue which arises inside the stem ground tissue, closer to the surface. Like the other dermal tissue (epidermis), it is a complex tissue. It includes three layers (starting from surface): phellemexternal layer of periderm, cork (cork), phellogencork cambium, lateral meristem making periderm (cork cambium) and phelloderminternal layer of periderm (Figure \(\PageIndex{7}\)). Phellem consists of large dead cells with secondary walls saturated with suberin, and is the main, thickest component of periderm. Phellogen is a lateral meristem, like cambium; it often arises fragmentarily (and also temporarily) and does not cover the whole stem under-surface. But when phellem starts to grow, all peripheral tissues (like epidermis) will be separated from water transport and eventually die. Phellogen makes phellem towards the surface, and phelloderm towards the next layer (phloem). Phelloderm is a minute tissue, and does not play significant role in the periderm.
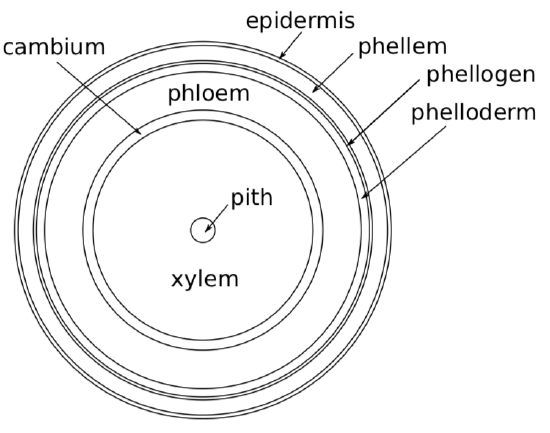
In older plants, phellogen arises deeper, sometimes inside phloem and separates outer layers of phloem from vascular cylinder. All this mixture of tissues (phellogen, phellem, phelloderm, epidermis and upper layers of phloem) considered as a bark.
Absorption Tissues
Poikilohydric plants that do not save water plants do not save water and they can survive even complete desiccation because their cells will hibernate. An example of a poikilohydric plants would be mosses. Homoiohydric plants that save water plants (which are majority of plants\(_2\)), however, do save water. They try to support the water content and do not survive complete desiccation. An example of a homoiohydric plant would be any “typical” plant, saying, corn. Somehow similar traits are comparable in poikilothermic animals, such as reptiles, and homoiothermic animals, such as birds and mammals, except in reference to body heat rather than water conservation.
Absorption tissues are always simple, primary tissues. Most important of them is rhizodermis(rhizoderm), or root hairs, which originates from protoderm (proto-epidermis), but its lifespan is much shorter than of epidermis. There are other absorption tissues, for example, velamen, which originates from the root cortex and consists of large, empty, easy to get wet dead cells.
Other Tissues
Secretory tissues spread across the plant body, concentrating in leaves and young stems. These tissues may secrete latex, volatile oils, mucus and other chemicals. Its functions can be attraction or dis-attraction, communication or defense, and many others.
In addition to tissues, plant body may contain idioblasts, cells which are quite dissimilar from surrounding cells. Idioblasts used for accumulation of unusual (and possibly dangerous) compounds like myrosinase, protein splitting glucosinolates into sugars and toxic isothiocyanate (mustard oil). We use mustard oil as a spice but for the plant, it works like a binary chemical weapon against insect herbivores: when myrosinase-containing idioblasts are damaged, mustard oil kills damaging insects. Among plants, the whole order Brassicales from rosids is capable to produce myrosinase, examples are different cabbages (Brassica spp.), papaya (Carica), horseradish tree (Moringa) and many others.