
1.31: Propagating plants and developing new plants
- Page ID
- 70350

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)
Ancient agriculture required plant propagation, making new plants. Plants don ' t live forever and many crops are annuals or are harvested less than a year after planting. Although the earliest agricultural efforts may not have involved intentionally propagating plants, fundamental to the development of agriculture was the acknowledgment that making new plants is an essential part of the process. The techniques used to propagate plant crops vary depending upon the crop. For most familiar crops it required storing some seed (i.e. not eating it) and planting it at some point in the future. For crops such as cereal grains and most legumes , where the seed is the part that is harvested and eaten, development of the ' store and plant ' technology was relatively straightforward and rapid. However crops such as tomato, squash and eggplant , where a fleshy fruit is harvested and the seeds are small and inconspicuous, required more technology: a recognition that seeds were present in the fruit and a method for processing of some of the fruit to obtain seeds for planting. Perennial plants do not require propagation each year, they could be maintained without propagation. But eventually they required some means of making new plants, especially if the agriculturalists are moving to a new area. For reasons discussed below, propagation of perennial plants often did not involve seeds but instead, asexual, means of propagation.
For most of its existence agriculture did not involve attempts to produce novel plants, plants with characteristics that distinguished them from their parents. However, the hope from the earliest onset of agriculture was that one might be able to improve the characteristics of the crop, producing crops that yielded more, tasted better, stored better, were easier to harvest, and were more tolerant of hardships, in particular, more tolerant of disease..Primitive farmers were happy when they found desirable novelties and also were able, knowingly or unknowingly, to gradually change the crops they grew. But they did not have the understanding and technology to intentionally create something novel. This has become possible over the last 100 years. Although an understanding of aspects of asexual reproduction was important for some crops, it was the recognition of the sexual process in pl ants, in particular the functioning of flowers and pollination , that has been critical to the development of the technologies of crop improvement (Figure 2). All of the major crops have been, and continue to be, intentionally modified. And this is one of the many factors that has changed, and made more complex, the business of agriculture. Where it was once basically controlled by a farmer who served both as a producer and salesman, it is now a vastly more complex system that depends on farmers, seed producers, researchers, fertilizer producers, equipment manufacturers, agricultural scientists, the government, and vast array of 'middle-men' .

TOPICS
- Asexual reproduction—propagation by cuttings
- Sexual reproduction by seeds and the importance of breeding system
- Hybrid seeds
- The green revolution
- Creating novel plants
- grafting
- generating variability
- genetic engineering
Asexual reproduction
Most woody plants do not 'breed true' , meaning that if you plant a seed from your favorite apple tree, or from a 'wild' tree growing in the woods, it is highly unlikely that the seed will grow into a tree comparable to the parent. In fact, the apples that you might harvest will probably be less than desirable. Similar patterns would be found for grapes and olives, two ancient crops that have long been propagated not from seed but instead asexually. The easiest technique is 'rooting' , cutting a stem and placing it in moist soil in hopes that it will sprout roots (see the discussion adventitious roots in Chapter 8). A similar procedure can be done with roots, inducing them to produce new shoots. Since the middle of the 20th century an artificial plant hormone has been used to promote adventitious growth in shoots/roots that might not do so otherwise.
Obviously, asexual propagation does not produce novel material, it o nly perpetuates existing ones. It is important in a large number of vegetable crops including banana, pineapple, potato, sweet potatoes, cassava and many more. It also is the most common means of propagating of most ornamental flowering plants started in the greenhouse (e.g. geranium, begonia). For some of these plants (e.g. banana, naval oranges and some ornamental flowers) asexual reproduction is essential because the plant is a sterile hybrid and cannot produce viable seeds. For other species, e.g. potato (Figure 3), pineapple (Figure 4) and many others asexual reproduction is simply an easier means of propagation, eliminating the seedling stage which is sometimes more sensitive to conditions than cuttings are. A disadvantage of asexual propagation is that the clones are genetically uniform and thus crops are more susceptible to widespread failure (e.g. in late blight of potato). Another problem is that viral diseases are transmitted in cuttings but not through seeds.


Sexual propagation —seeds and the significance of breeding system

Propagation by seeds is probably the most familiar process to most of us. For annual species grown as crops, propagation only requires the discipline to store seeds for the next season. Unlike asexual reproduction, sexual reproduction can result in variation. Variation has positive and negative effects on farming. On the one hand it allows for crop improvement and the production of novel plants (see below). On the other hand crop uniformity is generally helpful to agriculture because it makes cultural practices (e.g. planting and harvesting) more easily handled. Significant to crop variability is the breeding system of the crop. If the crop has a closed breeding system (Chapter 17) as the result of being apomictic or from having a bisexual flower and being self fertile, then offspring are likely to be the same as their parents and a crop may show little variation. Additionally, if a novel plant does appear, a closed breeding system makes it easier for a farmer to perpetuate plants with that specific feature. For instance, an important characteristic for cereal grains is to have 'non-shattering' heads, meaning inflorescences that do not shed their seeds, holding them on the plant and making harvesting easier. With a closed breeding system, seeds from non-shattering heads are likely to pass that feature on to their offspring since they may have no sexual process at all (apomixis) or are most likely to breed with themselves. In contrast, plants with a more open breeding system are more likely to be variable, with offspring that don 't all look alike and don' t necessarily look like their parent. And when a plant with favorable characteristics is found, it is more difficult to perpetuate these features through time. In fact, it is likely that these features will disappear quickly, from being 'washed out' by breeding with individuals that do not possess the feature. It is also significant that population size and reproductive isolation influence variability. A population of plants with an open breeding system may be very uniform if the population is small and reproductively isolated from other populations of the same plant, as can be the case for crop species.
Hybrid seed
Even before an understanding of the basis of genetics was developed the phenomenon of 'inbreeding depression' , had been noted: the decline in vigor in populations that are continually inbreeding. The degree to which this happens varies among species. It is probably not surprising that inbreeding depression is not a generally a problem for species with closed breeding systems. But for plants with open breeding systems, inbreeding can cause substantial reductions in crop yield. This was recognized by farmers and was part of the reason that seed companies, who took steps to avoid this problem, developed a clientele. Early in the 20th century plant breeders realized that although inbreed lines show reduced vigor, a 'hybrid cross' between two inbreed lines, produced plants that were more vigorous than either of the parental lines before they had been inbreed. Central to the production of hybrid seed is the need to control who breeds with whom. The technique requires that multiple generations bred only with close relatives, something that is easily accomplished by having populations isolated from each other. The next step is more difficult: ensuring that the inbreed lines NOT breed within the population but instead breed with individuals of another inbreed line. This was first done with corn, a plant with separate male and female flowers that are located in different parts of the plant. Crossing of inbreed lines can be accomplished by removing the male flowers (detasseling) from one inbreed line, thereby making these plants solely female and ensuring that if any seed is produced the male parent has come from a separate plant, usually from a second inbreed line that was planted nearby. Hybrid corn was first developed in the 1930's and became the dominant seed source by 1950, with yields increasing dramatically.
Large scale production of pure hybrid seed is nearly impossible in plants with perfect flowers (both male and female structures in the same flower). However, botanists discovered that it is possible to make plants with perfect flowers become unisexual. In the late 18th century workers noticed that not all pollen was viable and the ability to produce viable pollen sometimes often showed maternal inheritance. Later work indicated that there were cytoplasmic (i.e. non-nuclear) factors, inherited maternally (in plastids), that influence the production of viable pollen. Thus, geneticists were able to produce plants that were 'male sterile' , not able to produce viable pollen. Having plants that were unisexual (female) made the crosses that were needed to produce hybrid seed much more feasible. Hybrid seed production was further enhanced when 'restorer genes' were found that would restore the ability to make viable pollen. This allowed breeders to two cross two lines, one of which was unisexual, yet have the resulting hybrid produce flowers that are fully functional. Because of cytoplasmic male sterility the number of plants for which hybrid seed could be produced increased dramatically. Another means to the same end is the use of 'chemical hybridizing agents' , chemicals that make plants male (usually) sterile.
A negative consequence of planting hybrid seed is that seed cannot be stored year to year. Although the hybrid plant is vigorous, offspring of the hybrid plants are much less vigorous and also more variable. Growers utilizing hybrid seed need to purchase seed each year or switch to using open pollinated seeds instead of hybrid seed.
The Green Revolution
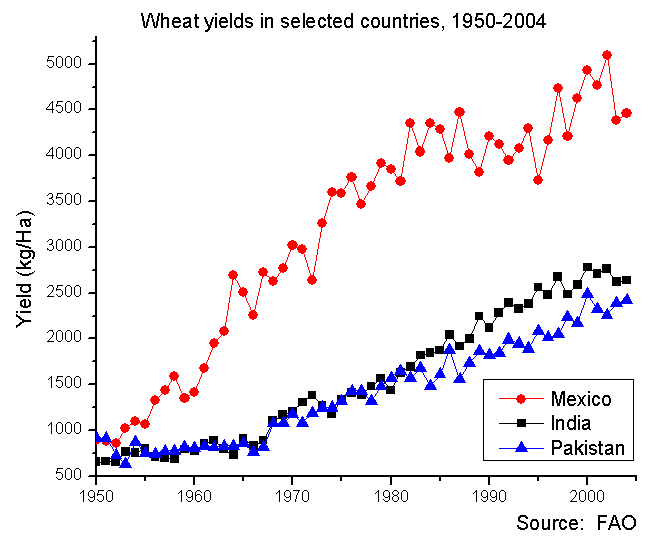
In the late 1960 's several scientists warned of an inevitable global famine, based in part on the assumption that crop yields could not be improved. Fortunately agricultural research, with a series of innovations that became known as the ' green revolution ' was able to drastically increase yield (Figure 6) and the global famine did not occur, although local famines continue to be a problem, usually the result of politics and war. Central to the green revolution were the efforts of Norman Borlaug, both as an agricultural researcher who developed high yielding varieties and as administrator who worked extensively to have new agricultural practices accepted by countries including Mexico, India and Pakistan. Central to the green revolution was hybrid seed and the development of new high yielding varieties of wheat and rice. Surprisingly, these varieties were actually dwarf plants. This had one direct effect: the problem of ' lodging' (plants being knocked over, usually as the result of wind) was lessened because the plants were shorter. But the new varieties also had a number of other features that helped to increase yield: increased seed production per plant, enhanced disease resistance (especially against rusts) and greater tolerance of drought or excessive rain. Also significant was that the new varieties would respond with increased yield to increased levels of fertilization. As a result of the new varieties and the new agricultural practices, crop yields more than doubled.
How can crops be improved? And who does the work of crop development?
Crop development requires careful observations and recognition of individuals with favorable traits. By the end of the 19th century the importance of generating variability was increasingly recognized as being significant to crop development. However, in general, farmers desire crop uniformity because it makes growing and harvesting easier and their focus is on growing crops not developing crops. During the 19th century, seeds were increasingly provided by the sources other than the farmer storing seeds or exchanging seeds with neighbors, and crop improvement was increasingly out of farmers'hands. During the latter half of the 19th century the federal government provided seed to anyone who asked for it. And, at the end of the century, land grant universities were charged with improving agriculture and this included developing improved varieties of crops. During this time period, private commercial seed companies became increasingly important as both a source of seeds and as a venue for improving crop quality.
Creating new plants using asexual methods of propagation
Although asexual reproduction is cloning, simply perpetuating what already exists, it has often been important in crop innovation. Novel plants are sometimes produced spontaneously by mutations of the seed or in branches of existing plants, and asexual reproduction of these mutants allows them to become new crops. This is what happened with navel oranges (Chapter 28), McIntosh apples (and many other apple varieties), poinsettia (Chapter 30), and a large number of ornamental plants. Cloning is also important as a means of perpetuating plants (generally hybrids) that have been created naturally (without human intervention) or artificially (with human intervention) that are unable to reproduce sexually.

Related to asexual reproduction is grafting, cutting a branch or bud off one plant and attaching it to a different plant, producing a novel organism, a chimera. There are a variety of reasons why this practice might be desirable, but one obvious one is in order to combine the favorable traits of two individuals into one individual, e.g. a good root system with a good shoot system. This is well represented by wine grape propagation. Stems from a European species are grafted on to root stocks from a North American species. The North American species produces inferior grapes for making wine but its root system can withstand attack from the insect pest Phylloxera. The root system of the European species is highly susceptible to Phylloxera but produces grapes of superior quality for wine. Phylloxera was mistakenly introduced from North America to Europe in the early 1800's and devastated European wine production. It was revived when workers were able to plant grapes composed of European shoots grafted on to North American rootstocks. Most fruit trees (apple, peach, plum) are similarly constructed with rootstocks that are vigorous and disease resistant grafted to shoots that produce desirable fruits.
Creating novel plants using the sexual process
Crossing with relatives
The relatives might be ancestral varieties that the crop originated from or species closely related to the crop. Technically, a crop species should should not hybridize with other species (i.e. it is an isolated gene pool, see Chapter 17) but sporadic seed production does occur. Although successful crosses may be infrequent, as long as there are some viable offspring, these can be back-crossed to the original crop (i.e. cross the hybrid plants with plants from the parental population) with significant introduction of novel traits (i.e. variability) that may include desirable features. Occasionally, workers are successful crossing with more distant relatives (different genera, even different families). Part of this surprising possibility may be poor taxonomy (i.e. they actually are more closely related than depicted in the taxonomy) but apparently sometimes crosses can happen between plants that are not that closely related.
Polyploidy
For a cross to be successful the parents need to have the same chromosome number and the chromosomes of the two parents need to be similar enough that they can pair during meiosis. When hybrids between different species are produced they are generally sterile because of the inability to pair chromosomes during meiosis. Although the hybrid is sterile it is often possible to propagate it asexually , i.e. to clone the hybrid, . A number of ornamental plants are perpetuated this way. It also is what has occurred with banana, the plant that is cultivated is a sterile hybrid, unable to produce seed but thiis one of the features that make the commercial banana desirable! Another 'solution' to the problem of hybrid sterility is polyploidy, increasing the chromosome number. This sometimes happens without human involvement (see discussion of the evolution of wheat in Chapter 28) but in the last 100 years workers have developed techniques to promote polyploidy following hybridization, thereby creating a new species with characteristics of both parental lines.
Treatment with mutagens
A common means of generating variability, and perhaps producing favorable traits that can be selected for, is to treat the seeds with a chemical mutagen or radiation. Although most of the treated seeds do not survive or have unfavorable features, usually some seeds survive that may have desirable features. These plants can be crossed with existing varieties in hopes of introducing favorable features to the crop. The technique is also used with asexual propagation. The original ruby red grapefruit appeared as a sport on a normal grapefruit tree. Irradiation of branches of the original ruby red has produced the even redder varieties Rio Red and Star Ruby.
Genetically modified organisms
A much more focused technique of combining traits from different organisms is the production of 'GMO' s, genetically modified organisms; the phrase should be non-sensical to any trained biologist because all organisms are genetically modified, that is what evolution is about. But if we focus only on agricultural organisms the term has come to distinguish 'normal' agricultural organisms, all of which have a long and substantial history of genetic modification by humans from 'abnormal' agricultural organisms, ones produced using the relatively recent techniques of molecular biology that allow for much more focused genetic modification, with genes manipulated in various ways including: turned on, turned off, duplicated, removed and, in particular, moved from one organism to another. One famous example is 'golden rice' , rice to which additional genes, genes that would cause the rice produce beta-carotene, a vitamin D precursor, into the endosperm of the seed. A second example is 'Bt' corn, which has a gene derived from the bacterium B acillus thuringiensis , that produces a protein that is toxic to some insects, in particular the corn borer moth. Consequently 'Bt-corn' can avoid predation by corn borers. Interestingly, the bacterial toxins had been used by organic farmers, being mass produced by culturing B. thuringiensis , extracting the toxins and then spraying them on corn plants. A final example is 'Roundup ready' soybeans. Roundup is a commonly used herbicide that is effective because it blocks an important synthetic pathway. Plant molecular biologists were able to introduce genes into several crops, including soybean and cotton, that greatly reduced the the toxicity of roundup to these crops, making them 'roundup ready' . The advantage of modern molecular techniques is specificity: plants are modified in very specific ways. In contrast, developing new crops by hybridization or by chemical or radiational means will have multiple effects — yet the breeder is only paying attention to a small number of characters they are hoping to modify.
GMO 's have been vigorously opposed by a variety of groups for a variety of reasons. This opposition has decreased substantially in the last five years, partly because many of the problems that were cited have not appeared even though GMO plants have been in wide use over the last fifteen years. Any large-scale production of crops with novel characteristics will have possible risks but the GMO' s are probably one of the more benign techniques to generate variability. Consequently the production and use of GMO s is likely to increase in the coming years.