
4.6.4: Germination
- Page ID
- 32044

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Learning Objectives
- Identify the environmental factors that stimulate germination.
- Distinguish between epigeous and hypogeous germination.
- Compare germination in eudicots versus monocots.
Many mature seeds enter a period of inactivity, or extremely low metabolic activity: a process known as dormancy, which may last for months, years or even centuries. Dormancy helps keep seeds viable during unfavorable conditions. Germination occurs when the embryo, which is dormant within a mature seed, resumes growth upon a return to favorable conditions. The embryo becomes a young seedling that is no longer confined within the seed coat.
In many seeds, the presence of a thick seed coat can inhibit germination through several mechanisms: (1) the embryo may not be able to break through the thick seed coat; (2) the seed coat may contain chemicals inhibitors; and (3) the seed coat prevents the embryo from accessing water and oxygen. Dormancy is also maintained by the relative hormone concentrations in the embryo itself.
Environmental Requirements for Germination
The requirements for germination depend on the species. Common environmental requirements include light, the proper temperature, presence of oxygen, and presence of water. Seeds of small-seeded species usually require light as a germination cue. This ensures the seeds only germinate at or near the soil surface (where the light is greatest). If they were to germinate too far underneath the surface, the developing seedling would not have enough food reserves to reach the sunlight. (Recall from 14.5 Dormancy that red light induces germination by converting the inactive form of phytochrome (Pr) to the active form (Pfr), which leads to the production of amylase. This enzyme breaks down the limited food reserves in the seed, facilitating germination.)
Not only do some species require a specific temperature to germinate, but they may also require a prolonged cold period prior to germination. In this case, cold conditions gradually break down a chemical germination inhibitor. This mechanism prevents seeds from germinating during an unseasonably warm spell in the autumn or winter in temperate climates. Similarly, plants growing in hot climates may have seeds that need a hot period in order to germinate, an adaptation to avoid germination in the hot, dry summers.
Water is always needed to allow vigorous metabolism to begin. Additionally, water can leach away inhibitors in the seed coat. This is especially common among desert annuals. Seeds that are dispersed by animals may need to pass through an animal digestive tract to remove inhibitors prior to germination. Similarly, some species require mechanical abrasion of the seed coat, which could be achieved by water dispersal. Other species are fire adapted, requiring fire to break dormancy (Figure \(\PageIndex{1}\)).
The Mechanism of Germination
The first step in germination and starts with the uptake of water, also known as imbibition. After imbibition, enzymes are activated that start to break down starch into sugars consumed by embryo. The first indication that germination has begun is a swelling in the radicle.
Depending on seed size, the time taken for a seedling to emerge may vary. Species with large seeds have enough food reserves to germinate deep below ground, and still extend their epicotyl all the way to the soil surface while the seedlings of small-seeded species emerge more quickly (and can only germinate close to the surface of the soil).
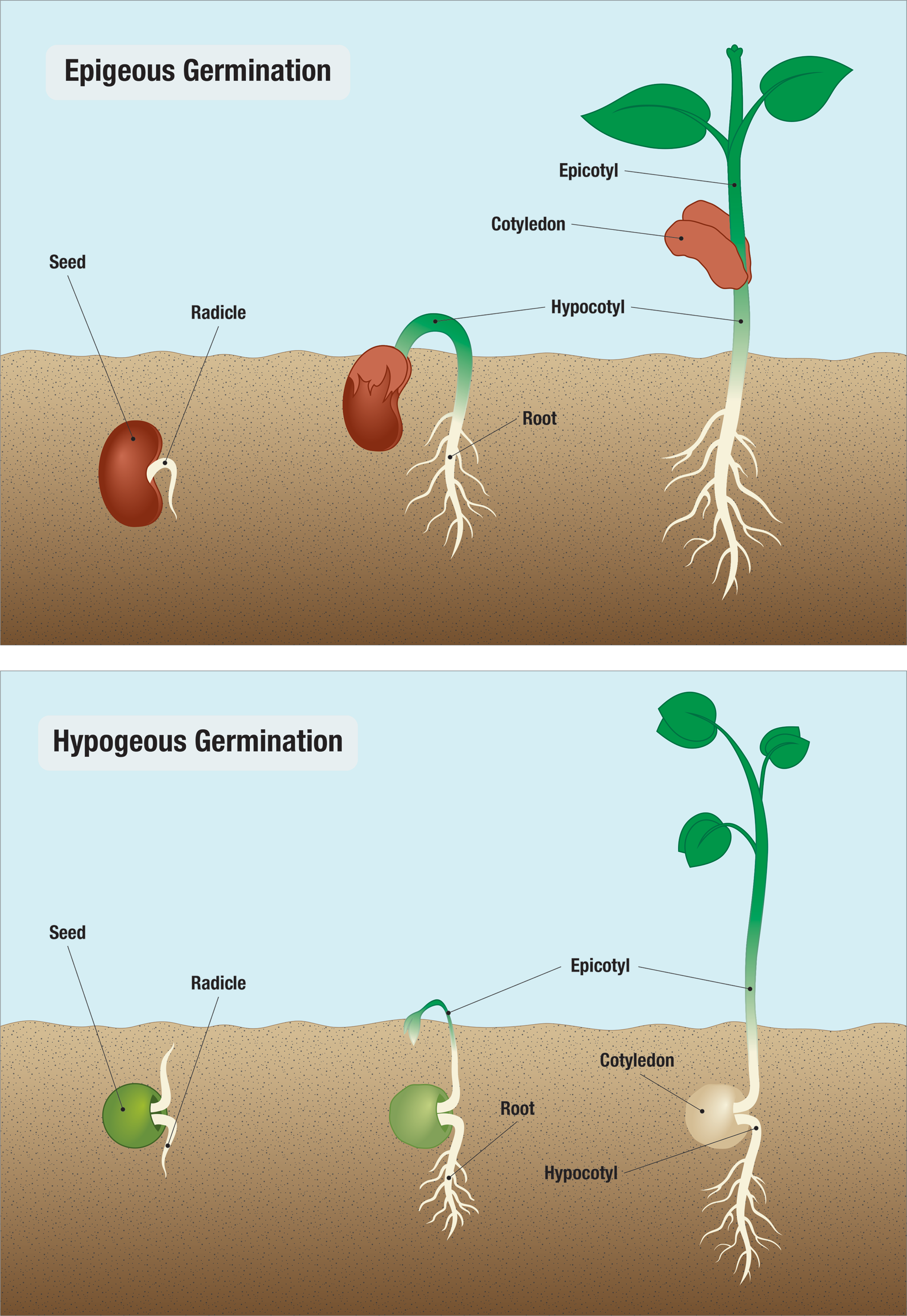
During epigeous germination, the hypocotyl elongates, and the cotyledons extend above ground. During hypogeous germination, the epicotyl elongates, and the cotyledon(s) remain belowground (Figure \(\PageIndex{2}\)). Some species (like beans and onions) have epigeous germination while others (like peas and corn) have hypogeous germination. In many epigeous species, the cotyledons not only transfer their food stores to the developing plant but also turn green and make more food by photosynthesis until they drop off.
Germination in Eudicots
Upon germination in eudicot seeds, the radicle emerges from the seed coat while the seed is still buried in the soil.
For epigeous eudicots (like beans), the hypocotyl is shaped like a hook with the plumule pointing downwards. This shape is called the plumule hook, and it persists as long as germination proceeds in the dark. Therefore, as the hypocotyl pushes through the tough and abrasive soil, the plumule is protected from damage. Additionally, the two cotyledons additionally protect the from mechanical damage. Upon exposure to light, the hypocotyl hook straightens out, the young foliage leaves face the sun and expand, and the epicotyl elongates (Figure \(\PageIndex{3}\)).


In hypogeous eudicots (like peas), the epicotyl rather than the hypocotyl forms a hook, and the cotyledons and hypocotyl thus remain underground. When the epicotyl emerges from the soil, the young foliage leaves expand. The epicotyl continues to elongate (Figure \(\PageIndex{4}\)).

The radicle continues to grown downwards and ultimately produces the tap root. Lateral roots then branch off to all sides, producing the typical eudicot tap root system.
Germination in Monocots
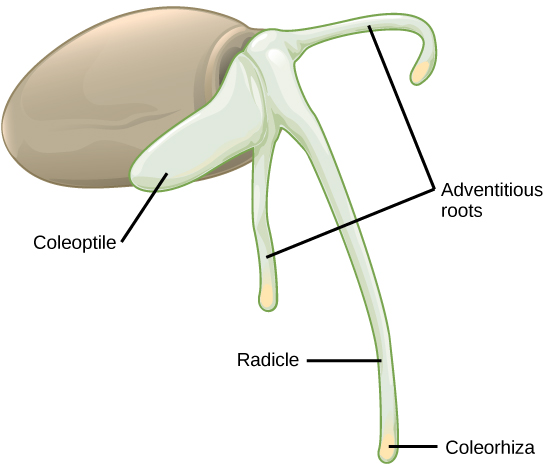
As the seed germinates, the radicle emerges and forms the first root. In epigeous monocots (such as onion), the single cotyledon will bend, forming a hook and emerge before the coleoptile (Figure \(\PageIndex{5}\)). In hypogeous monocots (such as corn), the cotyledon remains belowground, and the coleoptile emerges first. In either case, once the coleoptile has exited the soil and is exposed to light, it stops growing. The first leaf of the plumule then pieces the coleoptile (Figure \(\PageIndex{6}\)), and additional leaves expand and unfold. At the other end of the embryonic axis, the first root soon dies while adventitious roots (roots that arise directly from the shoot system) emerge from the base of the stem (Figure \(\PageIndex{7}\)). This gives the monocot a fibrous root system.


Attributions
Curated and authored by Melissa Ha using the following sources:
- 16.4B Germination of Seeds from Biology by John W. Kimball (CC-BY)
- 32.2 Pollination and Fertilization from Biology 2e by OpenStax (licensed CC-BY). Access for free at openstax.org.
- 7.5 Origin of the Seed from Introduction to Botany by Alexey Shipunov (public domain)