
4.5.1.2.2: Stomatal Opening and Closure
- Page ID
- 32036

Learning Objectives
- Relate the pattern of cell wall thickening in guard cells to their function.
- Explain the mechanism by which blue light triggers stomatal opening.
- Explain the mechanism by which water stress, signaled by abscisic acid, triggers stomatal closure.
Regulation of transpiration is achieved primarily through the opening and closing of stomata on the leaf surface. Stomata are surrounded by two specialized cells called guard cells (Figure \(\PageIndex{1}\)). Stomata must open to allow the gas exchange of carbon dioxide and oxygen for efficient photosynthesis (see Photorespiration), and light thus typically triggers stomatal opening. When stomata are open, however, water vapor is lost to the external environment, increasing the rate of transpiration. Therefore, plants must maintain a balance between gas exchange and water loss. Water stress, high temperatures, and high carbon dioxide concentration causes stomata to close.
Stomatal Opening
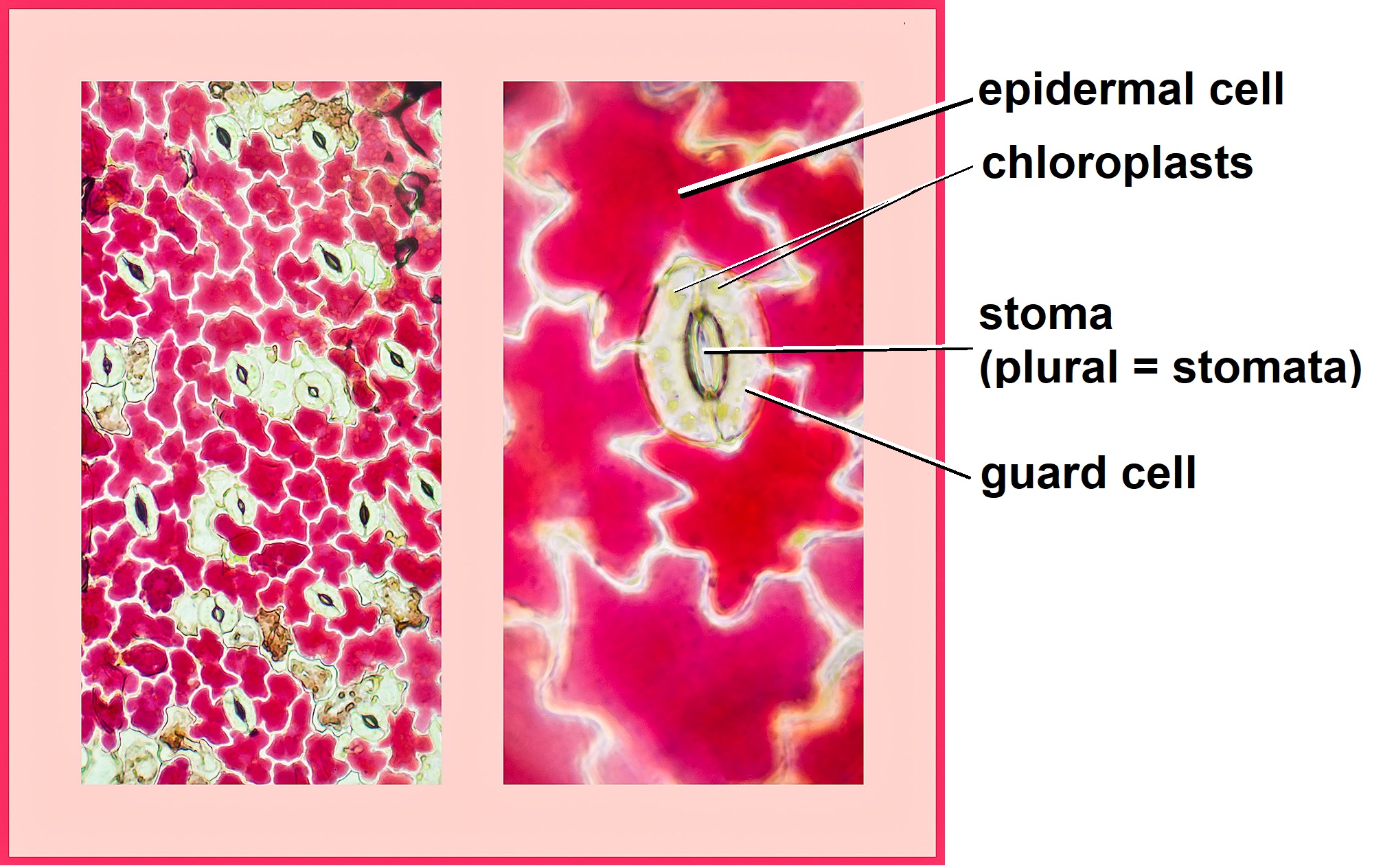
Guard cell walls are radially thickened such that the thickenings are concentrated around the stoma (plural: stomata; Figure \(\PageIndex{2}\)). When turgor pressure increases in guard cells, the cells swell. However, the thickened inner walls near the stoma cannot expand, so they curve to accommodate the expanding outer walls. The curving of the guard cells opens the stoma.

How does light cause stomata to open? Phototropins detect blue light, causing a proton pumps to export protons (H+). ATP, generated by the light reactions of photosynthesis, drives the pump. The cytosol usually more negative than the extracellular solution, and this difference in charge (membrane potential) increases as protons leave the cell. This increase in membrane potential is called hyperpolarization, and it causes potassium (K+) to move down its electrochemical gradient into the cytosol. Protons also move down their electrochemical gradient back into the cytosol, bringing chloride (Cl-) with them through symport channels. Meanwhile, starch is broken down, producing sucrose and malate. Nitrate (NO3-) also enters the cell. The solute potential resulting high concentrations of potassium, chloride, sucrose, malate, and nitrate in the cytosol drives the osmosis of water into the the guard cells. This increases turgor pressure, and the guard cells expand and bend, opening the stoma (Figure \(\PageIndex{3}\)).
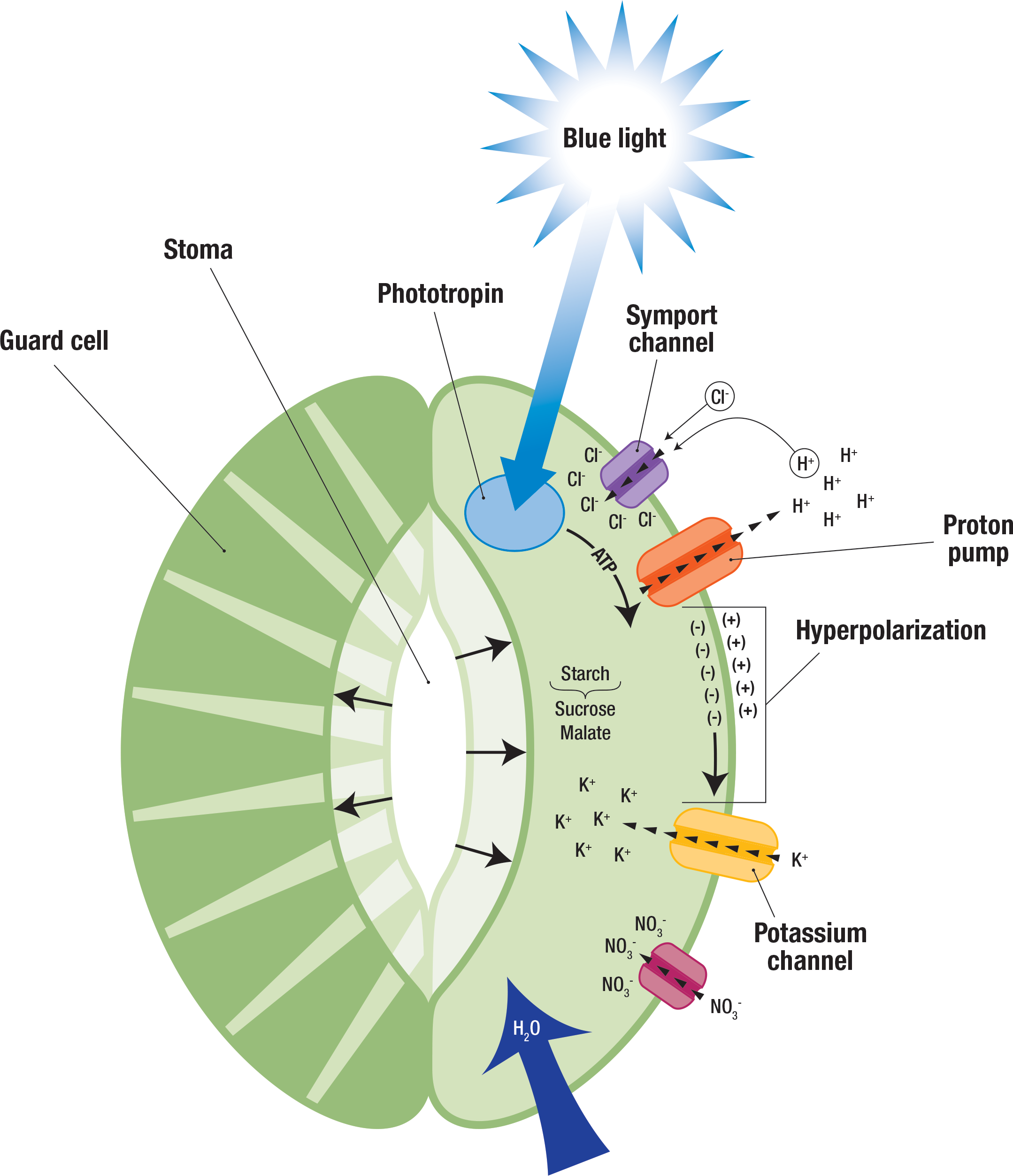
Table \(\PageIndex{1}\) illustrates how osmotic pressure (which results in turgor pressure) increases with light availability during the day. When the osmotic pressure of the guard cells became greater than that of the surrounding cells, the stomata opened. In the evening, when the osmotic pressure of the guard cells dropped to nearly that of the surrounding cells, the stomata closed.
| Time | Osmotic Pressure (MPa) |
|---|---|
| 7 A.M. | 1.46 |
| 11 A.M. | 3.14 |
| 5 P.M. | 1.88 |
| 12 Midnight | 1.32 |
Stomatal Closure
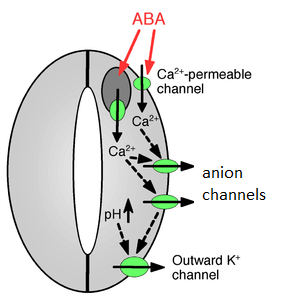
When water is low, roots synthesize abscisic acid (ABA), which is transported through the xylem to the leaves. There, abscisic acid causes calcium channels to open. Calcium (Ca2+) opens anion channels, and malate, chloride, and nitrate exit the cell. The membrane potential decreases (the difference in charge across the membrane becomes less pronounced) as anions leave the cell. Potassium exits the cell in response to this decrease in membrane potential (called depolarization). The loss of these solutes in the cytosol results in water leaving the cell and a decrease in turgor pressure. The guard cells regain their original shape, and the stoma closes (Figure \(\PageIndex{4}\)).
Attributions
Curated and authored by Melissa Ha using the following sources:
- 16.2D Gas Exchange from Biology by John. W. Kimball (licensed CC-BY)
- 30.2 Stems and 30.5 Transport of Water and Solutes in Plants from Biology 2e by OpenStax (licensed CC-BY). Access for free at openstax.org.