
3.4.2: Internal Leaf Structure
- Page ID
- 37052

Learning Objectives
- Describe the microscope internal structure of leaves, including the epidermis, mesophyll, and vascular bundles.
- Compare the adaptations of mesophytic, hydrophytic, and xerophytic leaves.
- Identify the unique features of pine and corn leaves.
- Compare the structures of sun and shade leaves.
Tissue Organization in Leaves
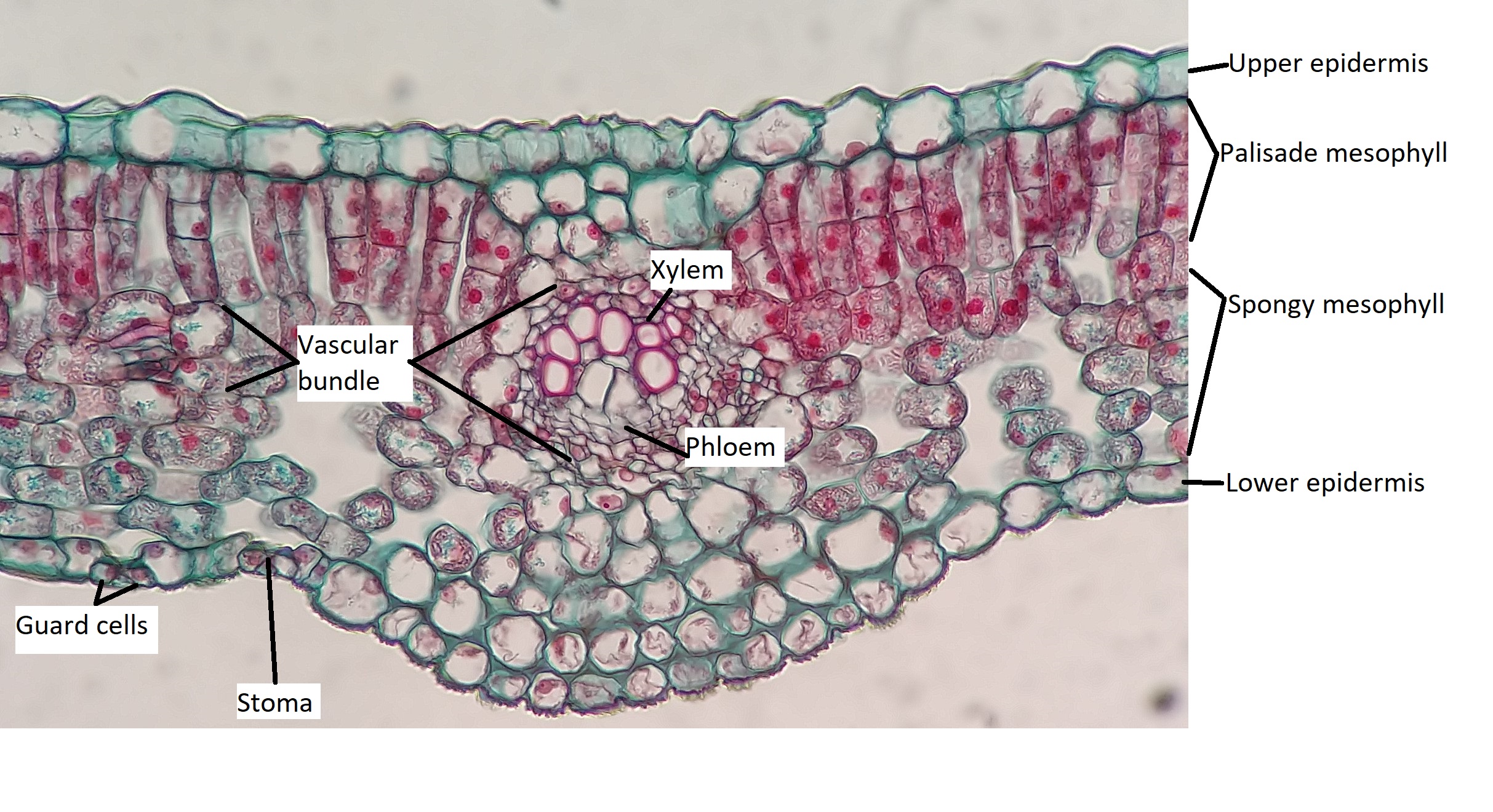
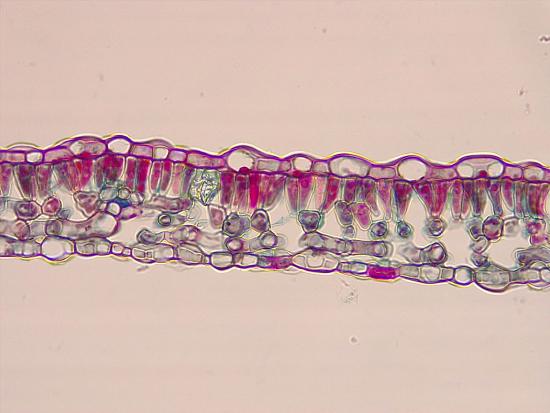
All three tissue types are represented in leaves. The epidermis represents the dermal tissue, the mesophyll that fills the leaf is ground tissue, and the vascular bundles that form the leaf veins represent vascular tissue (Figure \(\PageIndex{1}\)). These three tissues will be discussed using a eudicot leaf that is adapted to a moderate amount of water (mesophytic leaf). Variations in leaf structure are discussed later on this page.
Epidermis
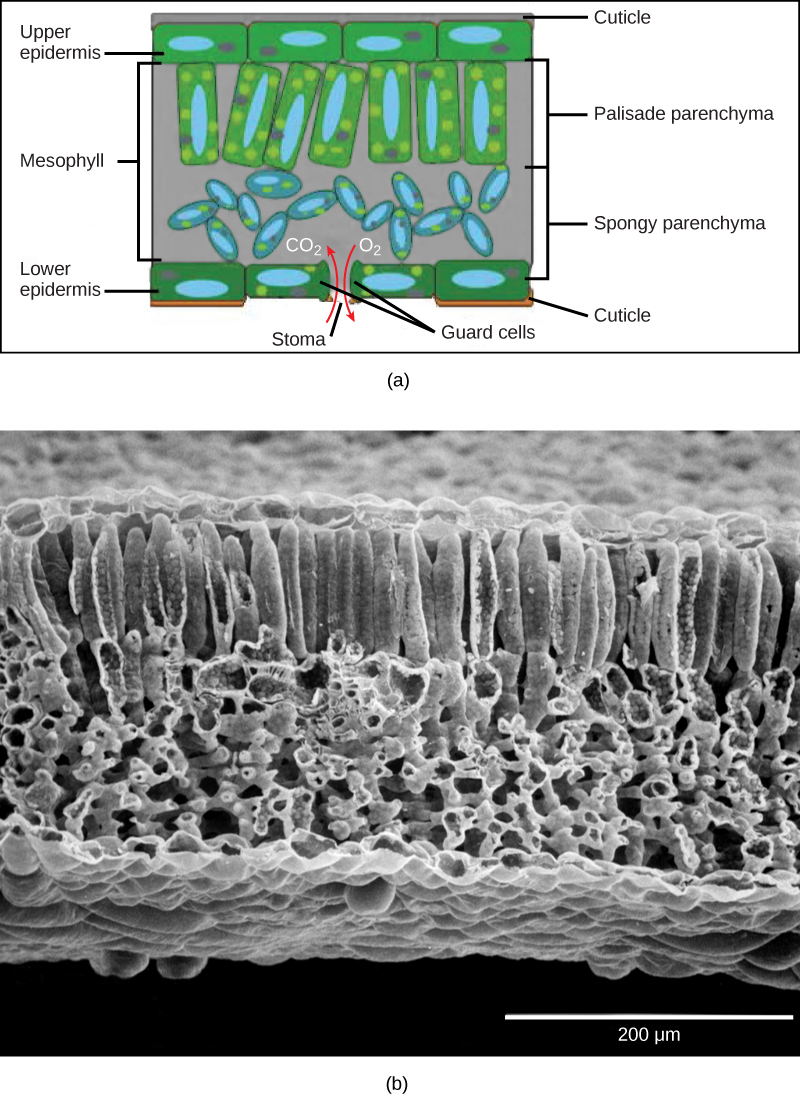
The outermost layer of the leaf is the epidermis; it is present on both sides of the leaf and is called the upper and lower epidermis, respectively. Botanists call the upper side the adaxial surface (or adaxis) and the lower side the abaxial surface (or abaxis). The epidermis helps in the regulation of gas exchange. It contains stomata (singular = stoma; Figure \(\PageIndex{2}\)), openings through which the exchange of gases takes place. Two guard cells surround each stoma, regulating its opening and closing, and the guard cells are sometimes flanked by subsidiary cells. Guard cells are the only epidermal cells to contain chloroplasts. In most cases, the lower epidermis contains more stomata than the upper epidermis because the bottom of the leaf is cooler and less prone to water loss.

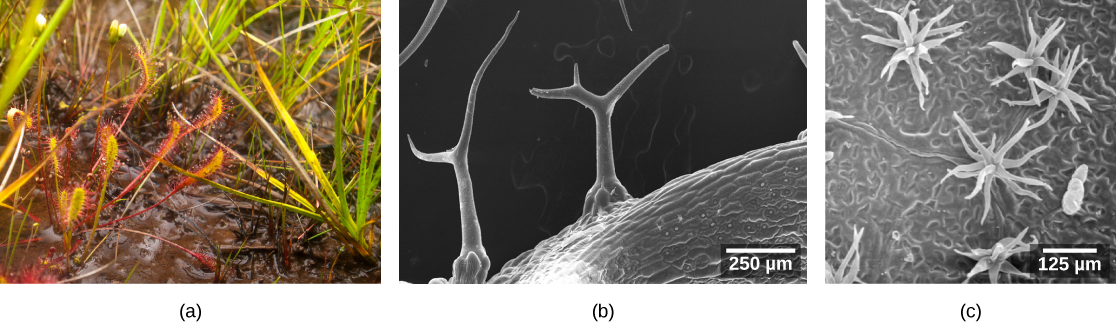
The epidermis is usually one cell layer thick; however, in plants that grow in very hot or very cold conditions, the epidermis may be several layers thick to protect against excessive water loss from transpiration. A protective layer called the cuticle covers surface of the epidermal cells (Figure \(\PageIndex{3}\)). The cuticle is rich in lignin (which lends some rigidity) and waxes (which function in waterproofing). The cuticle reduces the rate of water loss from the leaf surface. Other leaves may have small hairs (trichomes) on the leaf surface. Trichomes help to deter herbivory by restricting insect movements, or by storing toxic or bad-tasting compounds; they can also reduce water loss by blocking air flow across the leaf surface (Figure \(\PageIndex{4}\)). For this reason, trichomes (like stomata) are frequently denser on the lower side of the leaf.

Mesophyll
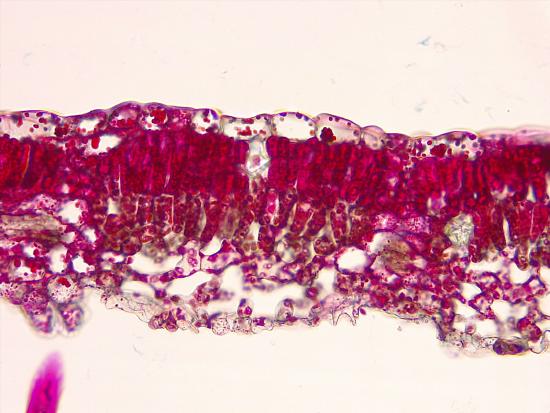
Below the epidermis are layers of cells known as the mesophyll, or “middle leaf.” Mesophyll cells contain many chloroplasts and specialize in photosynthesis. The mesophyll of most leaves typically contains two arrangements of parenchyma cells: the palisade parenchyma and spongy parenchyma (Figure \(\PageIndex{5}\)). The palisade parenchyma (also called the palisade mesophyll) has column-shaped and may be present in one, two, or three layers. The palisade cells specialize in capturing incoming sunlight (including slanted sun rays), rotating chloroplasts to the top of the leaf and then allowing them to regenerate by cycling them toward the leaf's center. They also decrease the intensity of sunlight for the spongy mesophyll. Although palisade cells may appear tightly packed in a cross section because there are many rows of cells behind those in the foreground, there is actually ample space (intercellular air spaces) between them. Below the palisade parenchyma are seemingly loosely arranged cells of an irregular shape. These are the cells of the spongy parenchyma (or spongy mesophyll). The intercellular air spaces found between mesophyll cells facilitate gaseous exchange.
Vascular Bundles (Veins)
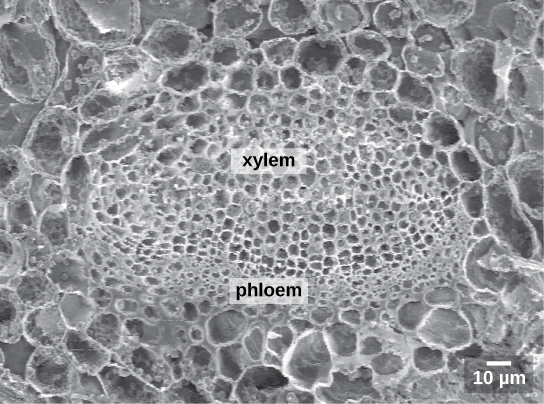
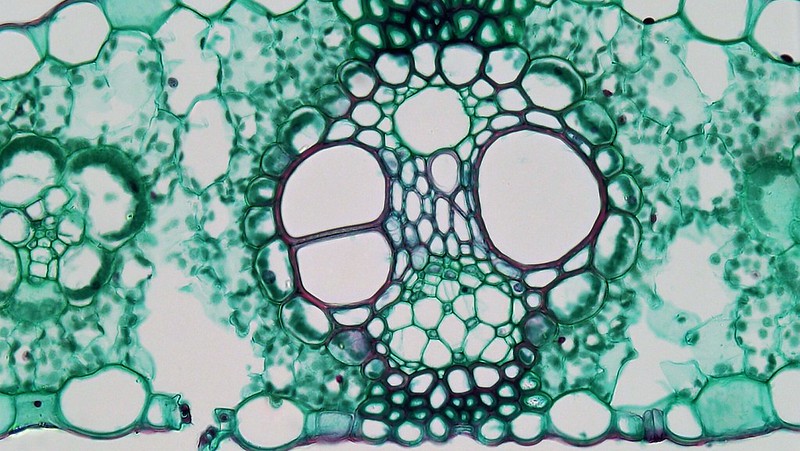
Like the stem, the leaf contains vascular bundles composed of xylem and phloem (Figure \(\PageIndex{6-7}\)). When a typical stem vascular bundle (which has xylem internal to the phloem) enters the leaf, xylem usually faces upwards, whereas phloem faces downwards. The conducting cells of the xylem (tracheids and vessel elements) transport water and minerals to the leaves. The sieve-tube elements of the phloem transports the photosynthetic products from the leaf to the other parts of the plant. The phloem is typically supported by a cluster of fibers (sclerenchyma) that increase structural support for the veins. A single vascular bundle, no matter how large or small, always contains both xylem and phloem tissues.

Leaf Adaptations
The broad, flat shape of most leaves increases surface area relative to volume, which helps it capture sunlight; however this also provides more opportunity for water loss. The anatomy of a leaf has everything to do with achieving the balance between photosynthesis and water loss in the environment in which the plant grows. Plants that grow in moist areas can grow large, flat leaves to absorb sunlight like solar panels because sunlight is likely more limiting than water. Plants in dry areas must prevent water loss and adapt a variety of leaf shapes and orientations to accomplish the duel tasks of water retention and sunlight absorption. In general, leaves adapted to dry environments are small and thick with a much lower surface area-to-volume ratio.
In regards to water, there are three main types of plants: mesophytes, hydrophytes, and xerophytes. Mesophytes are typical plants which adapt to moderate amounts of water ("meso" means middle, and "phyte" means plant). Many familiar plants are mesophytes, such as lilac, Ranunculus (buttercup), roses, etc. Hydrophytes grow in water ("hydro" refers to water). Their leaf blades are frequently highly dissected (deeply lobed) to access gases dissolved in water, and their petioles and stems have air canals to supply underwater organs with gases. Hygrophytes (not discussed further) live in constantly wet environment, their leaves adapted to rapidly release water through the stomata. They sometimes even excrete of water drops through the leaf margins (guttation). Xerophytes are adapted to the scarce water ("xero" refers to dryness). Xerophytes are found in deserts and Mediterranean climates (such as in much of California), where summers are hot and dry. The leaves of mesophytes are called mesophytic, hydrophyte leaves are called hydrophytic, and so on. The structure of mesophytic leaves was already described (Figure \(\PageIndex{1}\)). Adaptaions in hydrophytic and xerophytic leaves and discussed below in more detail.
Hydrophytic Leaves
The structure of a hydrophytic leaf differs from a mesophytic leaf due to selective pressures in the environment -- water is plentiful, so the plant is more concerned with staying afloat and preventing herbivory. Hydrophytic leaves have a thin epidermal layer and the absence of stomata in the lower epidermis (Figure \(\PageIndex{8}\)). In the spongy mesophyll, there are large pockets where air can be trapped, helping the leaf float. This type of parenchyma tissue, specialized for trapping gases, is called aerenchyma. Sharp, branched sclereids (astrosclereids) traverse the mesophyll of a hydrophytic leaf. These provide the leaf structural support, as well as prevention of herbivory. Vascular tissue is somewhat reduced in hydrophytic leaves.


Xerophytic Leaves
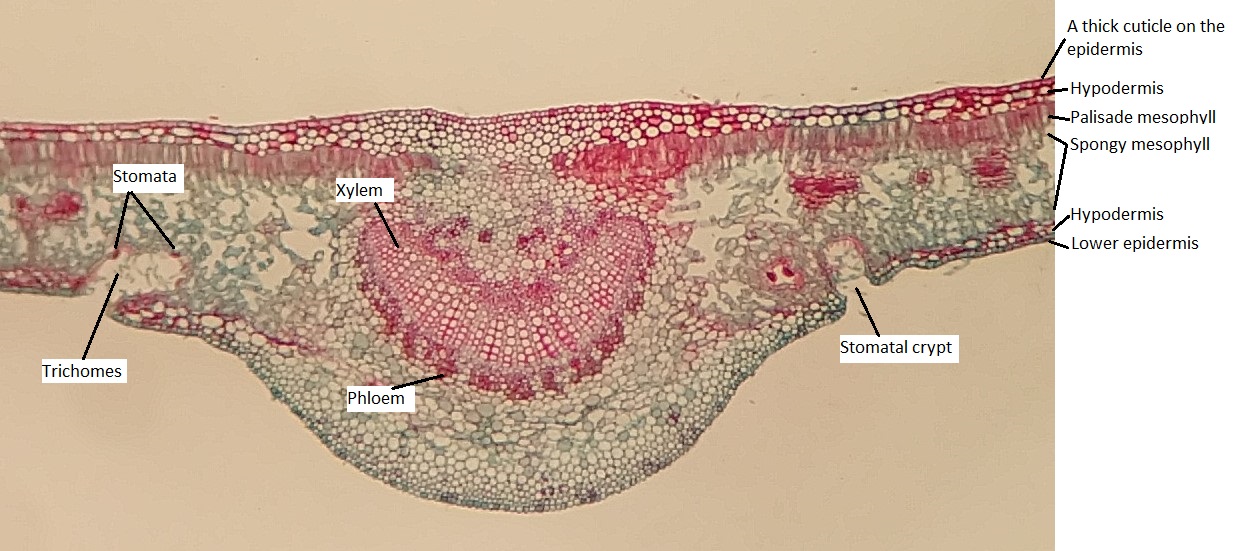
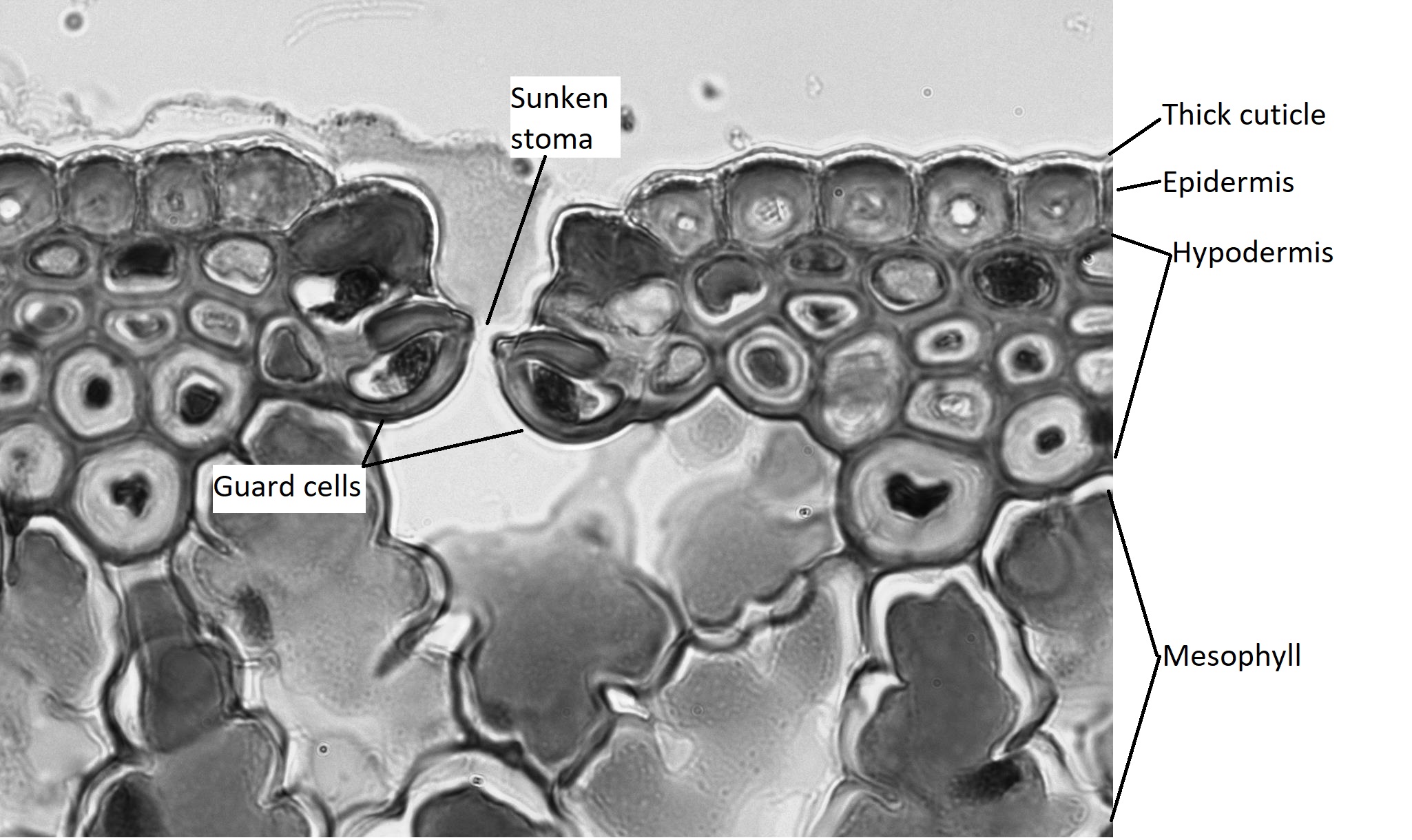
Xerophytic leaves (Figure \(\PageIndex{9}\)) have thick cuticles to limit water loss, especially on the upper epidermis (Figure \(\PageIndex{10}\)). Both the upper and lower epidermis consists of several layers (multiple epidermis). Sometimes the additional layers are called the hypodermis ("hypo" meaning under; "dermis" meaning skin). Depressions in the lower epidermis creates a pockets that are lined with trichomes, and the stomata are located at the base of these pockets (called stomatal crypts; figure \(\PageIndex{10}\)). The trichomes help capture evaporating moisture and maintain a relatively humid environment around the stomata. These stomatal crypts are located only on the underside of the leaves, where they experience less sun exposure and therefore less water loss. The upper epidermis is free from stomata.
Pine Leaves
Pines evolved during a period in Earth’s history when conditions were becoming increasingly dry, and pine needles have many adaptations to deal with these conditions. Many of these adaptations are similar the xerophytic leaves of some angiosperms (described above) because pines themselves are xerophytes.
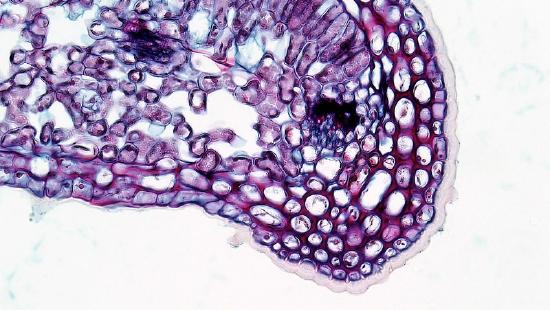
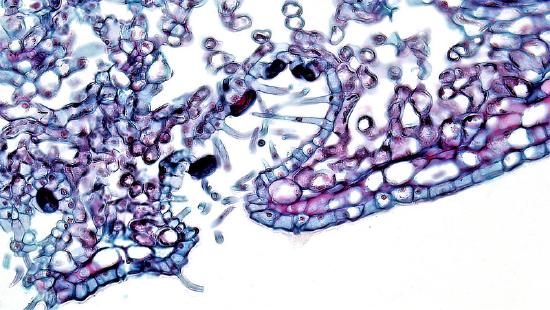
The epidermis of the leaf seems to be more than one cell layer thick (figure \(\PageIndex{11}\)). These subsequent layers of epidermis-like tissue under the single, outer layer of true epidermis are called the hypodermis , which offers a thicker barrier and helps prevent water loss. The epidermis itself is coated on the outside by a thick layer of wax called the cuticle. Because waxes are hydrophobic, this also helps prevent water loss through the epidermis. The stomata are typically sunken, occurring within the hypodermis instead of the epidermis. Sunken stomata create a pocket of air that is protected from the airflow across the leaf and can aid in maintaining a higher moisture content (figure \(\PageIndex{12}\)).
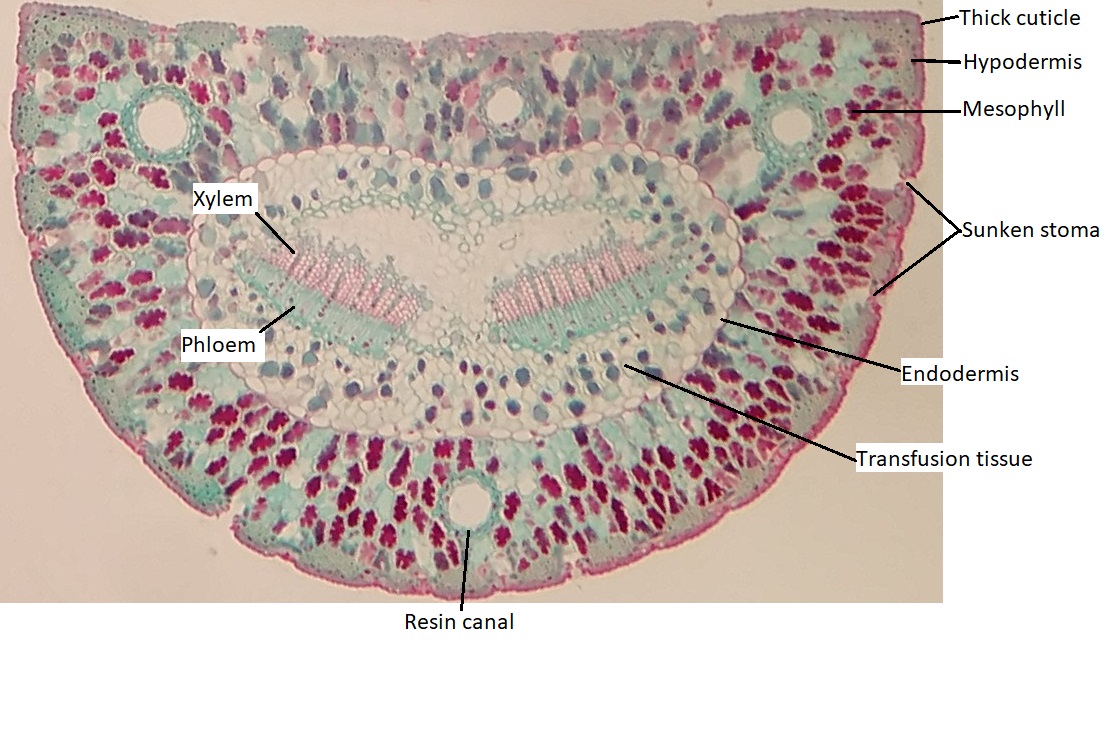
Within the mesophyll, there are several canals that appear as large, open circles in the cross section of the leaf. These are resin canals. The cells lining them secrete resin (the sticky stuff that coniferous trees exude, often called pitch), which contains compounds that are toxic to insects and bacteria. When pines evolved, not only was the Earth becoming drier, but insects were evolving and proliferating. These resin canals are not features that help the plant survive dry conditions, but they do help prevent herbivory. In addition to prevention of herbivory, resin can aid in closing wounds and preventing infection at wound sites.
There are two bundles of vascular tissue embedded within a region of cells called transfusion tissue, which functions in transporting materials to and from the mesophyll cells. The transfusion tissue and vascular bundles are surrounded by a distinct layer of cells called the endodermis. This is similar to the tissue of the same name in the root, but the cells are not impregnated with the water-repelling compound suberin.
Finally, the overall shape of the leaf allows for as little water loss as possible by decreasing the relative surface area, taking a rounder shape as opposed to a flatter one. This low surface area-to-volume ratio is characteristic of xerophytes.
Corn Leaves
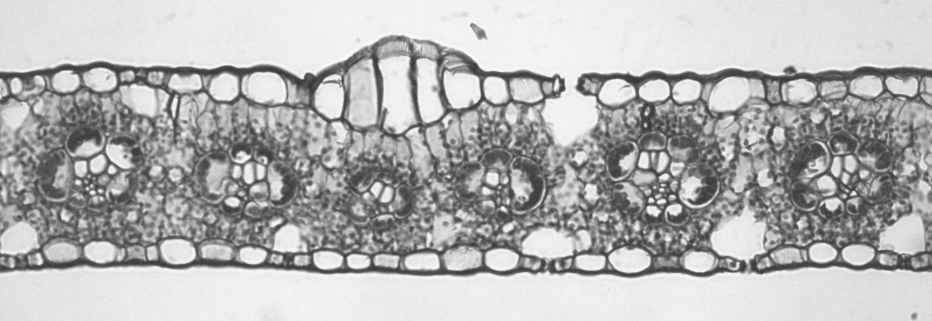
The model organism for monocots in botany is usually corn (Zea mays). In corn, there are approximately the same number of stomata on both the upper and lower epidermis. The mesophyll is not divided into two distinct types. The vascular bundles all face the same directly (appearing circular in cross section) because they run parallel to each other.
Corn is not necessarily a xerophyte, but it is adapted to deal with high temperatures. One of these adaptations, C4 type photosynthesis is discussed in Photorespiration and Photosynthetic Pathways and results in a cell arrangement called Kranz anatomy. The vascular bundles are surrounded by obviously inflated parenchyma cells that form a structure called a bundle sheath, and these are packed with chloroplasts (Figure \(\PageIndex{13}\)). (Bundle sheaths surround vascular bundles of other types of leaves as well, but the bundle sheath cells are much smaller). Mesophyll cells encircle the bundle sheath cells. In C4 photosynthesis, carbon dioxide is first gathered by the mesophyll cells and temporarily stored as a four-carbon sugar. This four-carbon sugar is transferred to the bundle sheath cells, where it is broken down to release carbon dioxide. It is in the bundle sheath cells where a process called the Calvin cycle, and glucose is ultimately produced. C4 photosynthesis concentrates carbon dioxide inside the bundle sheath cells, reducing the need to frequently open stomata for gas exchange. This helps conserve water.
When moisture is plentiful, the corn leaves are fully expanded and able to maximize photosynthesis. When moisture is limited, the leaves roll inward, limiting both moisture loss and photosynthetic capacity. This is accomplished by the presence of bulliform cells in the upper epidermis (Figure \(\PageIndex{14}\)). These clusters of enlarged cells are swollen with water when there is abundant water available. As the water content in the plant decreases, these cells shrivel, causing the upper epidermis to curl or fold inward at these points. This adaptation to sun exposure can be found in many other grasses, as well (corn is a member of the Poaceae, the grass family).
Sun and Shade Leaves
The light intensity experienced by a developing leaf influences its structure. Leaves that develop when consistently exposed to direct sunlight (sun leaves) thus differ from leaves exposed to low light intensities (shade leaves) in several ways (Figure \(\PageIndex{15}\)). Relative to shade leaves, sun leaves are smaller and thicker. This reduces surface area relative to volume, conserving water, which would otherwise be easily lost under bright sunlight and resultantly warmer temperatures. In contrast, the broad, thin shape of shade leaves helps capture sufficient light when light intensity is low. The thicker cuticle of sun leaves also limits water loss. They have more palisade parenchyma and more vascular tissue. Sun leaves can maintain a high photosynthetic rate at high light intensities, but shade leaves cannot.
Attributions
Curated and authored by Melissa Ha using the following sources:
- 30.2 Stems and 30.4 Leaves from Biology 2e by OpenStax (licensed CC-BY). Access for free at openstax.org.
- 5.3 The Leaf from Introduction to Botany by Alexey Shipunov (public domain).
- 9.2 Introduction to Leaf Anatomy and 9.3 Leaf Anatomy from Botany Lab Manual by Maria Morrow (licensed CC-BY)
- 13 Leaves from A Photographic Atlas for Botany by Maria Morrow (licensed CC-BY-NC).