
3.3.3: Secondary Stem
- Page ID
- 27718

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Learning Objectives
- Compare the origin and function of the vascular cambium and cork cambium.
- Define bark and distinguish between inner and outer bark.
- Explain the production of wood and relate this to annual rings.
- Distinguish between heartwood and sapwood.
- Distinguish between softwood and hardwood.
- Identify the external features of winter twigs.
Primary growth occurs as a stem increases in length as a result of cell division in the shoot apical meristem. Secondary growth is characterized by an increase in thickness or girth of the plant, and is caused by cell division secondary meristems. Herbaceous plants mostly undergo primary growth, with hardly any secondary growth or increase in thickness. Secondary growth or “wood” is noticeable in woody plants; it occurs in some eudicots, but occurs very rarely in monocots.
Secondary Meristems (Lateral Meristems)
Two secondary meristems (lateral meristems) are responsible for secondary growth: the vascular cambium and cork cambium (Figure \(\PageIndex{1}\)).
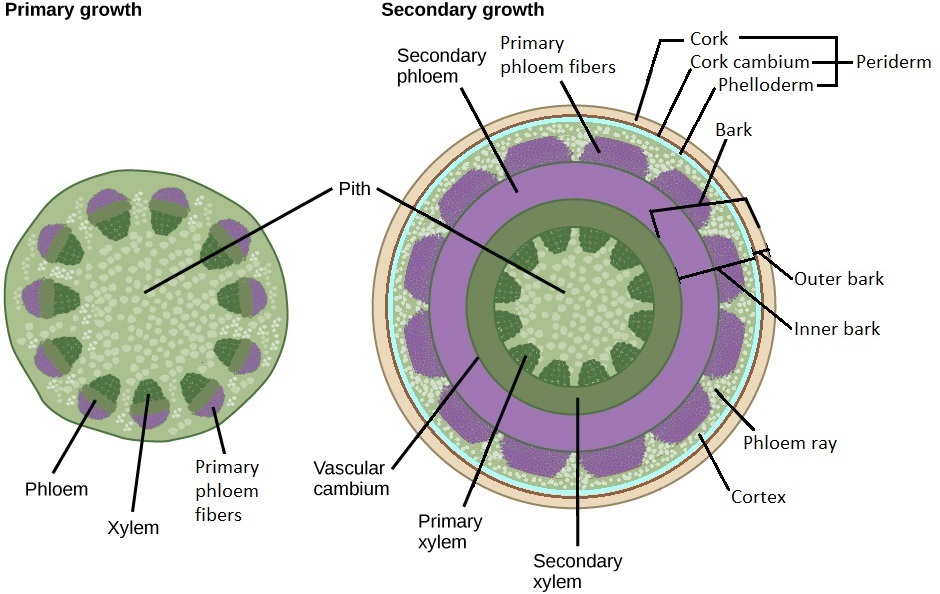
The vascular cambium produces secondary vascular tissue. The fusiform initials are the cells of the vascular cambium that divide to produce secondary xylem internally and secondary phloem externally. The ray initials are the cells of the vascular cambium that produce vascular rays (xylem rays and phloem rays). These are bands of parenchyma that are perpendicular to the concentric layers of xylem and phloem (Figure \(\PageIndex{2}\)). They function in storage, producing secondary compounds (molecules used by the plant that are not essential parts of metabolism), and transporting materials between the xylem and phloem. As the secondary stem thickens, the phloem rays thicken externally (becoming wedge-shaped) to accommodate the increasing diameter. While the vascular cambium is technically only a single layer cell layer, it looks similar to the layers of cells that surround it (that it recently divided to produce), and this entire region is sometimes called the vascular cambium as a result.

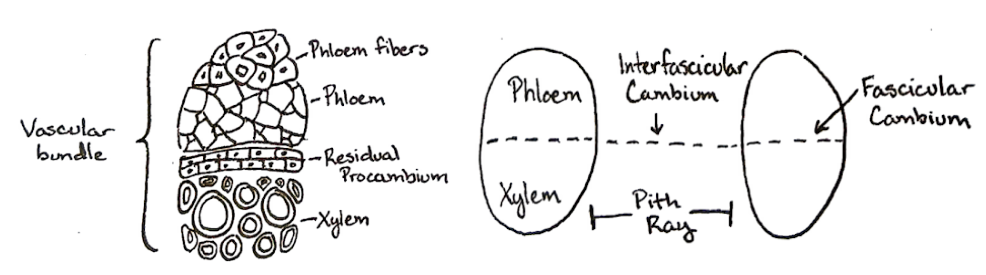
The vascular cambium arises from stem cells within and between the vascular bundles in some silenosteles and eusteles. Within vascular bundles, such stem cells (specifically, procambial cells) form the fascicular cambium. In the interfascicular regions between vascular bundles is interfascicular cambium (Figure \(\PageIndex{3}\)). The fascicular cambium and interfascicular cambium ultimately form the vascular cambium. In contrast, the vascular cambium in roots arises from the procambium and pericycle.
The cork cambium divides to produce phelloderm internally and cork externally. Together, the phelloderm, cork cambium, and cork form the periderm, the dermal tissue of the secondary plant body (figure \(\PageIndex{4}\)). The first cork cambium produced by a stem arises from the cortex, but subsequent cork cambia are produced by the parenchyma cells of the secondary phloem. (In contrast, the cork cambium arises from the pericycle in roots.)

Palm trees, which are monocots, do not have secondary meristems and true wood. Some thickening does occur in a palm but this happens at the base of the tree, as a result of adventitious roots growing. Palms may also have diffuse secondary growth which is division and enlargement of some parenchyma cells. These processes do not compensate the overall growth of plant, and palms frequently are thicker on the top than on the bottom. Another monocot, dragon blood tree (Dracaena), has anomalous secondary growth, which employs cambium but this cambium does not form the stable ring.
Periderm and Bark
At the end of the secondary stem's first year of growth, the periderm replaces the epidermis, but the cortex and pith are retained. In contrast, roots that undergo secondary growth do not have piths to begin with, and the cortex is lost during secondary growth. Like the epidermis, most of the periderm is not permeable to water vapor, carbon dioxide, and gaseous oxygen. This is due to the waxy suberin that fills the cork cells, which are dead at maturity. However, gas exchange with the environment is possible at lenticels, elevated regions of the periderm with many intercellular air spaces (Figure \(\PageIndex{5-6}\)). To produce lenticels, some cork cambium cells divide and grow much faster, which will finally break the periderm open. Woody stems do not do regular gas exchange as primary stems do by opening and closing stomata, but woody plants still have leaves with high densities of stomata to regulate gas exchange.


Bark consists of all of the tissue layers external to the vascular cambium. It protects the plant against physical damage and helps reduce water loss. In a one-year stem from inside to outside, this would be the secondary phloem, primary phloem fibers, cortex, phelloderm, cork cambium, and cork. The cork cambium divides the inner and outer bark. The inner bark is everything within the cork cambium. The outer bark is the cork cambium and everything external to it (Figure \(\PageIndex{7}\)).
A secondary stem ultimately produces multiple layers of periderm. The inner bark in an older stem thus consists of the newest secondary phloem and the newest phelloderm. Only the conducting phloem of the inner bark contains live cells and transports materials while the nonconducting phloem of the inner bark contains dead cells that are used for storage. As the secondary stem ages, the old layers of the secondary phloem are pushed externally and crushed, with the exception of the phloem fibers, which have thickened cell walls. The outer bark in an older stem would be the newest cork cambium, newest cork, and concentric layers of old phloem and old periderm. If the multiple periderms form perfect circles, the bark is smooth. More often, multiple periderm do not overlap evenly, resulting in rough bark with scales.

Wood (Secondary Xylem)
Wood consists of the secondary xylem produce by the vascular cambium (Figure \(\PageIndex{8}\)). In contrast to the phloem, old layers of secondary xylem are retained and are not easily crushed. However, the oldest secondary xylem (close to the center of the secondary stem) no longer conducts water. This is the heartwood, which stores various compounds and appears darker than the surrounding wood. To block the flow of water in the heartwood, plants use tylosesvessel element “stoppers”, which also help control winter functioning of vessels. A tylose forms when a cell wall of parenchyma grows into the tracheary element; they look like bubbles. The sapwood surrounds the heartwood, is lighter in color, and consists of the conducting xylem, which was more recently produced (Figure \(\PageIndex{7}\)).

In the spring of temperate regions, the vascular cambium produces wide tracheary elements (the conducting cells of the xylem, either vessel elements or tracheids). These transport large volumes of water, which is abundant due to spring rains. During the summer, the vascular cambium produces narrow tracheary elements as a result of lower water availability. In the winter, the vascular cambium's activity is low. It resumes the next spring by again producing the wide tracheary elements of early wood (spring wood), which distinctly contrast with the adjacent late wood (summer wood) from the previous year. Early wood appears lighter and is less dense than late wood. Each year of wood production is thus visible in a cross section of a woody stem because it consists of a light layer and a dark layer. These are called annual rings (tree rings; Figure \(\PageIndex{9-10}\)) and can be used to determine the age of a tree or branch through the study of dendrochronology. Furthermore, thick annual rings indicate wet years, and thin annual rings indicate dry years.


Some trees (like oaks, Quercus) have large vessel elements are found primarily in early wood; this pattern is known as ring porous (Figure \(\PageIndex{10}\)). Large vessel elements of other trees (like elm, Ulmus) occur more evenly in both early and late wood. This pattern is known as diffuse porous wood: with large vessel elements in both early and late wood. (Diffuse porous species still produce annual rings due to differences in tracheid size.) Trees growing in climates without well-expressed seasons, such as the tropical rainforest, will not make annual rings at all.
Hardwoods are produced by angiosperms and contain both vessel elements and tracheids (figure \(\PageIndex{10}\)). Softwoods are produced by conifer trees (in the gymnosperm phylum Coniferophyta) and contain only tracheids (Figure \(\PageIndex{11}\)). These terms are misnomers to an extent, however, because hardwoods are not always denser than softwoods. Xylem rays tend to occupy a greater volume in hardwoods relative to softwoods. Additionally, the arrangement of cells appears more disorderly in hardwoods due to the large size of vessel elements. Finally, softwoods contain resin ducts (Figure \(\PageIndex{11}\)), which contain a thick substance (resin) important in defense and response to injury.

Winter Twigs
Winter deciduous trees and shrubs in temperate regions become dormant in winter. The twigs of these species have the basic external features of a stem (axillary buds, nodes, etc.), but they are modified to facilitate dormancy in the winter and resumption of growth in the spring. At the end of a winter twig is the terminal bud, which contains a shoot apex surrounded by protective structures called bud scales. When the terminal bud resumes growth, the bud scales fall off and leave marks called terminal bud-scale scars. These form a ring around the twig, marking the winter of each year. Lateral buds are similar in structure to terminal buds, but they are found at each node. Just below the lateral buds are leaf scars, where the leaves were formerly attached. Within the leaf scars are bundle scars, marking leaf traces (consisting of vascular bundles) that moved from the stem to the leaf (Figure \(\PageIndex{12}\)).

Attributions
Curated and authored by Melissa Ha from the following sources:
- From 5.4: The Stem from Introduction to Botany by Alexey Shipunov (public domain)
- From 30.2 Stems from Biology 2e by OpenStax (licensed CC-BY)