
2.7.4: Angiosperm Families
- Page ID
- 27727

Learning Objectives
- Interpret a floral formula.
- Describe the characteristics used to identify each of the four largest angiosperm families.
Knowing who is related to whom in the plant world can provide important information. Often, closely related organisms will have similar life history traits, such as defense compounds or other secondary metabolites. Identifying which family an unknown plant belongs to might provide insight into potential interactions with other organisms (see Orchidaceae below), its capability to resist particular environmental conditions or stressors, its edibility or toxicity, or whether it produces compounds that might be used medicinally.
For example, plants like poison oak (Toxicodendron diversilobum) and poison ivy (Toxicodendron radicans) are in the family Anacardiaceae and produce an oil called urushiol. This oil causes a (sometimes severe) allergic reaction in many people, resulting in an itchy, blistering rash. Mango (Mangifera indica) and cashew (Anacardium occidentale). also belong to this family, but these are not known for giving itchy rashes. However, as someone who is quite allergic to poison oak, I unknowingly ate a mango with the skin on and broke out in a rash all around my mouth and lower face, just like how I react to touching poison oak. Indeed, cashews are just the seed of the cashew fruit. The rest of the fruit and seed coat are removed due to the urushiol content.
Additionally, while there are over 350,000 species of flowering plants, there are only around 400 families. Being able to quickly narrow an unknown plant to family is an essential skill in plant identification. So how can we tell who belongs to the same family among plants? For flowering plants, we can use floral formulas.
Floral Formulas
Since there are so many terms about flowers, and at the same time, flower structure and diversity always were of immense importance in botany, two specific ways were developed to make flower description more compact. First is a flower formula. This is an approach where every part of flower is designated with a specific letter, numbers of parts with digits, and some other features (whorls, fusion, position) with other signs.
- Symmetry: * means radial symmetry, while X means bilateral symmetry.
- Whorls: K is the calyx, C is the corolla, A is the androecium, and G is the gynoecium. The number that follows each letter represents the number of parts in that whorl. For the gynoecium, a line under the number indicates an inferior ovary, while a line above the number indicates a superior ovary.
- Fusion: In most representations, connation is indicated by circling the number, while adnation is indicated by drawing a line connecting the numbers of the fused whorls.
Here are a few examples of floral formulas, followed by their interpretation:
\(\ast K_{4}C_{4}A_{2+4}G_{\underline{(2)}}\): flower actinomorphic, with four sepals, four petals and six stamens in two whorls, ovary superior, with two fused carpels
\(\uparrow K_{(5)}[C_{(1,2,2)}A_{2,2}]G_{\underline{(2\times2)}}\): flower zygomorphic, with five fused sepals, five unequal fused petals, two-paired stamens attached to petals, superior ovary with two subdivided carpels
\(\ast K_{(5)}C_{(5)}[A_{5}G_{\underline{(3)}}]\): actinomorphic flower with five fused sepals and five fused petals, five stamens attached to pistil, ovary inferior, with three fused carpels
The following signs are used to enrich formulas:
PLUS “+” is used to show different whorls; minus “\(-\)” shows variation; “\(\vee\)” = “or”
BRACKETS “[]” and “()” show fusion. In most representations, connation is indicated by circling the fused whorl, while adnation is indicated by drawing a line underneath the formula connecting those whorls.
COMMA “,” shows inequality of flower parts in one whorl
MULTIPLICATION “\(\times\)” shows splitting
INFINITY “\(\infty\)” shows indefinite number of more than 12 parts
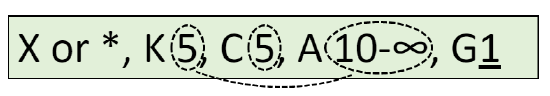
Flower diagram is a graphical way of flower description. This diagram is a kind of cross-section of the flower. Frequently, the structure of pistil is not shown on the diagram. Also, diagrams sometimes contain signs for the description of main stem (axis) and flower-related leaf (bract). The best way to show how to draw diagram is also graphical (Figure \(\PageIndex{1}\)); formula of the flower shown there is \(\ast K_{5}C_{5}A_{5}G_{\underline{(5)}}\).
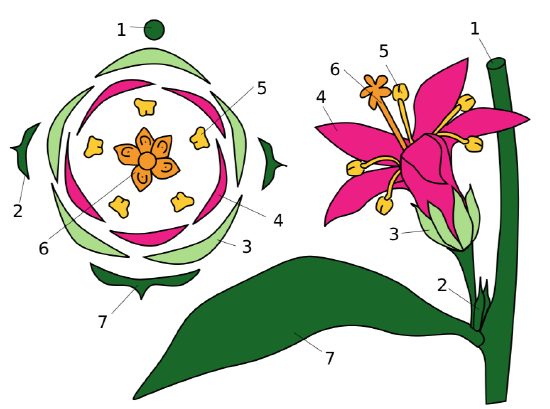
Review of Terms and Formula Designations
FLOWER PARTS occur in whorls in the following order—sepals, petals, stamens, pistils.
(The only exceptions are flowers of Eupomatia with stamens then perianth, Lacandonia with pistils then stamens, and some monocots like Triglochin, where stamens in several whorls connect with tepals.)
PEDICEL flower stem
RECEPTACLE base of flower where other parts attach
HYPANTHIUM cup-shaped receptacle (Figure \(\PageIndex{2}\))
PERIANTH = CALYX + COROLLA
SEPALS small and green, collectively called the CALYX, formula: K
PETALS often large and showy, collectively called the COROLLA, formula: C
TEPALS used when sepals and petals are not distinguishable, they form SIMPLE PERIANTH, formula: P
ANDROECIUM collective term for stamens: formula: A
STAMEN = FILAMENT + ANTHER
ANTHER structure containing pollen grains
FILAMENT structure connecting anther to receptacle
GYNOECIUM collective term for pistils/carpels, formula: G. Gynoecium can be composed of:
- A single CARPEL = simple PISTIL, this is MONOMERY
- Two or more fused CARPELS = compound PISTIL, this is SYNCARPY
- Two or more unfused CARPELS = two or more simple PISTILS, this is APOCARPY
(Note that variant #4, several compound pistils, does not exist in nature.)
To determine the number of CARPELS in a compound PISTIL, count LOCULES, points of placentation, number of STYLES, STIGMA and OVARY lobes.PISTIL Collective term for carpel(s). The terms CARPEL and PISTIL are equivalent when there is no fusion, if fusion occurs then you have 2 or more CARPELS united into one PISTIL.
CARPEL structure enclosing ovules, may correspond with locules or placentas
OVARY basal position of pistil where OVULES are located. The ovary develops into the fruit; OVULES develop into seeds after fertilization.
LOCULE chamber containg OVULES
PLACENTA place of attachment of OVULE(S) within ovary
STIGMA receptive surface for pollen
STYLE structure connecting ovary and stigma
FLOWER Floral unit with sterile, male and female zones
ACTINOMORPHIC FLOWER A flower having multiple planes of symmetry, also called radially symmetrical, formula: \(\ast\)
ZYGOMORPHIC FLOWER A flower having only one plane of symmetry, also called bilaterally symmetrical, formula: \(\uparrow\)
PERFECT FLOWER A flower having both sexes
MALE / FEMALE FLOWER A flower having one sex, formula: ♂ / ♀
MONOECIOUS PLANTS A plant with unisexual flowers with both sexes on the same plant
DIOECIOUS PLANTS A plant with unisexual flowers with one sex on each plant, in effect, male and female plants
SUPERIOR OVARY most of the flower is attached below the ovary, formula: \(G_{\underline{\dots}}\)
INFERIOR OVARY most of the flower is attached on the top of ovary, formula: \(G_{\overline{\dots}}\)
(Inferior ovary only corresponds with monomeric or syncarpous flowers.)
WHORL flower parts attached to one node
Major Families
This section will cover some of the largest families of angiosperms, including their floral formula and general characteristics.
Orchidaceae, the Orchid Family
Orchids are one of the most species-rich group of plants, containing over 28,000 species (Figure \(\PageIndex{1}\)). These plants tend to be tropical and epiphytic (growing on other plants). However, as can be assumed from their vast diversity, orchids can be found in many ecosystems and growing on a variety of substrates, including rocks! Their flowers are often highly modified, including long nectar spurs, hairy petals, and strange morphologies. Many orchids are fly pollinated. Follow this link to see observations of orchids from across the globe and in your region!
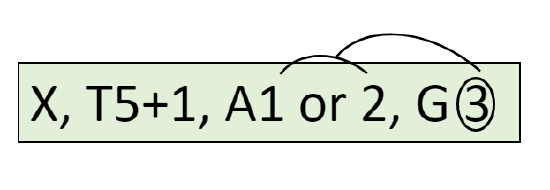
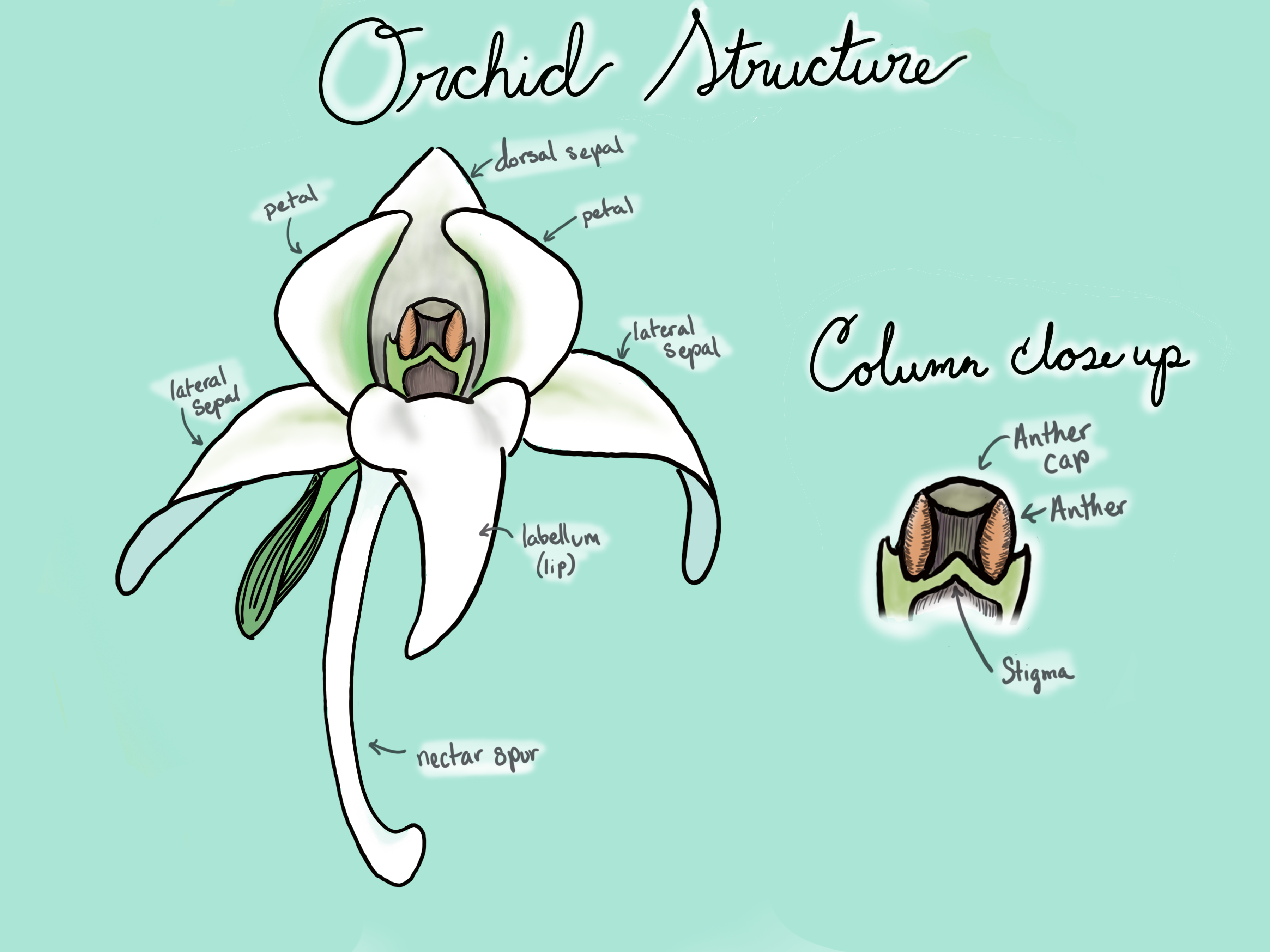


Orchids make a large number of tiny seeds, i.e. the tiny black specks in vanilla bean ice cream are the seeds of the orchid Vanilla plantifolia. An interesting, r-selected strategy with a twist: the seeds parasitize mycorrhizal fungi. Orchid seeds will not germinate if they do not have a fungal partner. As the fungal hyphae penetrate into the orchid cells, they formed coiled structures. The orchid feeds on the fungal hyphae until it has produced its first leaves and can photosynthesize on its own. At this point, the relationship can be shifted toward mutualism, with sugars transfered from plant to fungus. However, some orchids have lost the ability to make chlorophyll and instead, continue to feed from their fungal partner. These plants are referred to as mycoheterotrophs. The mycoheterotrophic orchid Rhizanthella gardneri undergoes its entire life cycle underground!
Asteraceae, the Aster Family or Composite Family
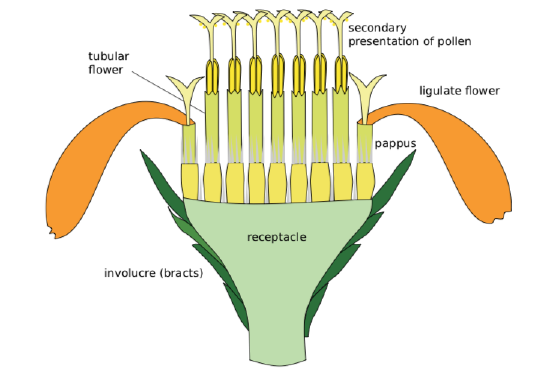
There are more than 32,000 accepted species in Asteraceae—a recent and dramatic increase in described species has sent them soaring past the orchids (for the time being). They have a cosmopolitan distribution, but are better represented in temperate and subtropical regions. This family is primarily herbaceous but does contain some woody species. Members of this family produce head inflorescences with one or two different types of florets that all attach to a common receptacle (Figure \(\PageIndex{4}\)). Disc florets have radial symmetry, while ligulate florets have bilateral symmetry (Figure \(\PageIndex{5}\)). Some inflorescences contain only disc florets (e.g. thistle), some only ligulate florets (e.g. dandelions), and others contain a combination of the two (e.g. daisy). The inflorescence is subtended by layers of bracts called phyllaries, forming an involucre.
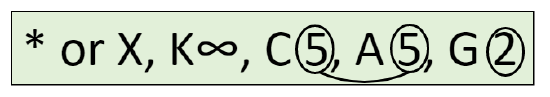
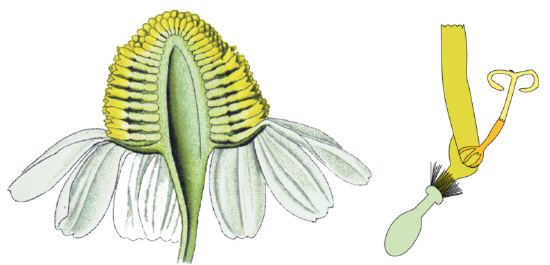
The calyx is reduced to hairs or bristles (pappus, see Figure \(\PageIndex{6-7}\)), petals are fused into a tube or ligula (with 5 or 3 teeth). The pollen is lifted up and distributed by the outer sides of the stigmas, called secondary pollen presentation (Figure \(\PageIndex{6}\)). Florets have inferior ovaries. The fruit is an achene and the mature seed has almost no endosperm.

Many plants withing the Asteraceae are used for oils, vegetables, ornamentals and medicinal plants are distributed in multiple subfamilies. The most commercially important of these are:
- Carduoideae: mostly tubular flowers
-Centaurea—knapweed
-Cynara—artichoke
-Carthamus–safflower
- Cichorioideae: mostly 5-toothed ligulate (pseudo-ligulate) flowers + lacticifers with latex
-Taraxacum—dandelion
-Lactuca—lettuce
- Asteroideae: tubular + 3-toothed ligulate flowers
-Helianthus—sunflower (BTW, “canola”, or Brassica napus from Cruciferae is the second main source of vegetable oil)
-Artemisia—sagebrush
-Tagetes—marigold and lots of other ornamentals
Fabaceae, the Legume Family
With around 19,000 species, Fabaceae (Figure \(\PageIndex{8}\)) is the third largest angiosperm family after orchids and asters. This family is widely distributed throughout the world, but does particularly well in the tropics. These plants form root nodules with nitrogen-fixing bacteria. Leaves are alternate, pinnately compound (once or twice), with stipules. Plants in this family have legumes as fruits (one locule dehiscent along a single suture).
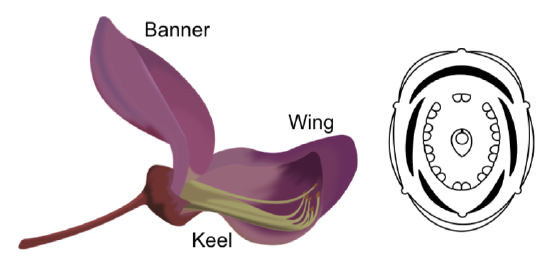
There are three subfamilies (Caesalpinioideae, Mimosoideae, Papilionoideae) with distinct characteristics. In Papilionoideae the petals are mostly free, unequal and have special names: banner, keel and wing (Figure \(\PageIndex{9}\)). In Mimosoideae, they fuse and form a tube with radial symmetry. There are usually 10 stamens with 9 fused and one free; in Mimosoideae, stamens are numerous. There is a single carpel in the gynoecium, meaning the ovary will have a single locule.
Some representatives of Fabaceae include:
- Mimosoideae: stamens numerous, petals connected
-Acacia—dominant tree of African and Australian savannas, often with phyllodes
-Mimosa—sensitive plant
- Papilionoideae: stamens 9+1, petals mostly free; this subfamily contains many extremely important food plants with high protein value
-Glycine—soybean
-Arachis—peanut with self-buried fruits
-Phaseolus—bean
-Pisum—pea
Poaceae, the Grass Family
Grasses are monocots. There are approximately 12,000 species of grasses widely distributed throughout the world, but most genera concentrate in the tropics. Grasses tend to be wind dispersed and prefer dry, sunny places. They often form turf (tussocks)—compact structures where old grass stems, rhizomes, roots, and soil parts are intermixed. Grasses form grasslands—specific ecological communities widely represented on Earth (for example, North American prairies are grasslands). Grasses are also important components of other ecosystems (e.g. wetlands). Stems of grasses are usually hollow and round. The leaves have sheathing bases.
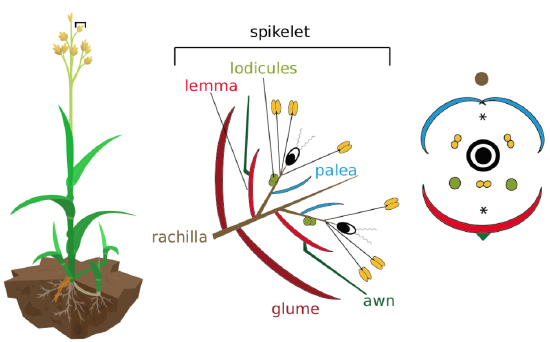
Grass florets are reduced, wind-pollinated, usually bisexual (Figure \(\PageIndex{10}\)), and form complicated spikelets. Each spikelet bears two glumes; each flower has lemma and palea scales (Figure \(\PageIndex{11}\)). The perianth is reduced to lodicules. Stamens from 6 to 1 (most often 3), with large anthers. The fruit of grasses is a caryopsis, what we commonly refer to as a grain. Grasses are incredibly important commercially as food sources, construction, biofuels, and components of many products: rice, corn, wheat, sugarcane, barley, rye, sorghum, and bamboo are all grasses. The study of grasses is called agrostology--it can be both aggravating and glumey.
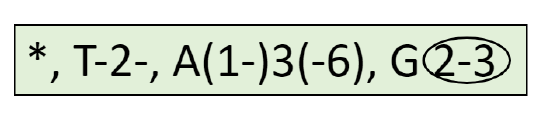
Attribution
Curated and authored by Maria Morrow using the following sources:
- 19.1.5 Diversity and Evolutionary Relationships of the Plants from Biology by John. W. Kimball (licensed CC-BY)
- 8.3 Three plant families you wanted to know but were too afraid to ask from Introduction to Botany by Alexey Shipunov (public domain)
- Angiosperm Families presentation by Melissa Ha, (CC BY-NC)