
2.6.2.2: Conifers and Gnetophytes
- Page ID
- 37014

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Use morphological features and life history traits to distinguish conifers from other plants.
- Connect the adaptations of conifers to dry and/or cold environments.
- Identify structures and phases in the Pinus life cycle; know their ploidy.
- Use morphological features and life history traits to distinguish gnetophytes from other plants.
- Describe the traits gnetophytes share with angiosperms.
Though the gnetophytes have been difficult to place, phylogenetically, recent genetic studies place them as sister to the Pinaceae (pine family, emerging from within the conifers. See this open-access paper for recent genetic work on the evolutionary relationships between gymnosperms.
Conifers
Conifers are the most species-rich lineage of gymnosperms. From the fossil record, we think there were over 20,000 species of conifers. However, their diversity declined with the dinosaurs. Currently, there are around 600 extant species. These amazing plants represent some of the oldest, tallest, and most massive organisms on the planet. Though currently low in diversity, these amazing plants make up 30% of Earth’s forests.
Conifers are the most widely known and economically important among gymnosperms. Conifers include the largest and the oldest of all living organisms. One redwood (Sequoia sempervirens) growing in California is almost 400 feet (122 meters) high. Bristlecone pines (Pinus longaeva, Figure \(\PageIndex{1}\)) growing in the mountains of eastern California some are more than 5,000 years old. Giant sequioas (Sequiadendron giganteum) Most of them are temperate evergreen trees, but some are deciduous, such as larch (Larix) and the dawn redwood (Metasequoia). The stem has a large amount of xylem, a small cork, and minute pith. Seeds are distributed by wind and animals.
.jpg?revision=1&size=bestfit&width=550&height=550)
Note: The Pinaceae is currently the largest family of conifers, so many of the examples for this group of gymnosperms will be from the type genus Pinus (pines).
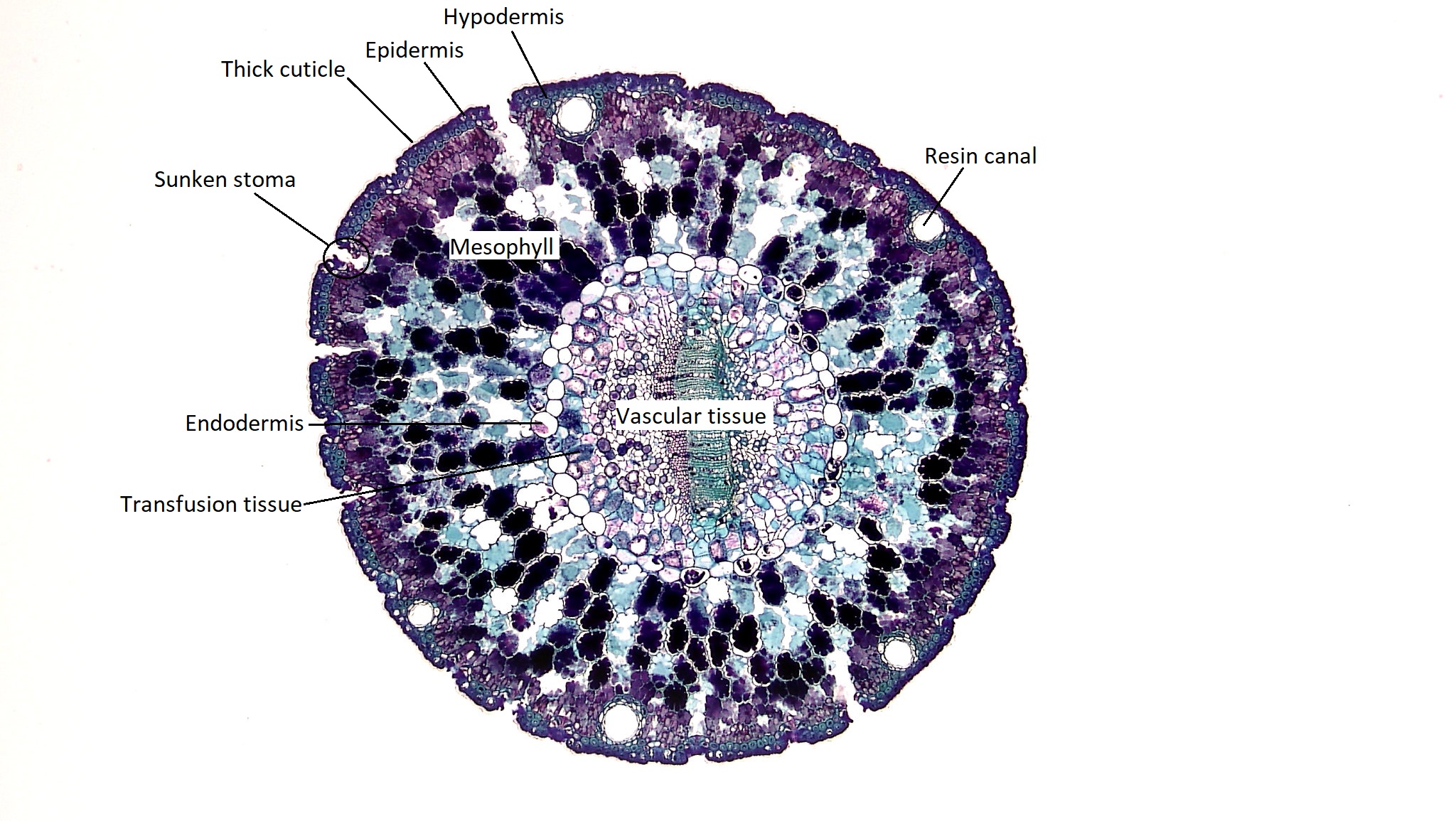
Xerophytic Leaves in Conifers
Xerophytic leaves are adapted to withstand drought conditions. In conifers, we see a wide range of xerophytic leaves with different morphologies that can be shaped by their local environment. Consider the leaves of the coast redwood and the giant sequoia (Figure \(\PageIndex{2-3}\)). Though these two trees belong to different genera--Sequoia and Sequoiadendron, respectively--they are sister taxa. However, the coast redwood has adapted to life on the coast, where the giant sequoia has evolved in inland, higher elevation forests with much more extreme climatic conditions. How can this be seen in the structure of their leaves?


 Figure \(\PageIndex{3}\): These giant sequoia leaves were retrieved from a tree planted in a coastal environment, so the differences here represent evolutionary history, alone. Much like the coast redwood, these shiny leaves have a thick cuticle. The big difference to note is the thickness of these leaves, maintaining a much lower surface area to volume ratio than the coast redwood. The second thing to note is the orientation of the leaves. They are held much more closely together, reducing exposure to the wider environment. Two stomata are indicated in the image, though there are many. They are present on both adaxial and abaxial leaf surfaces. Photo by Maria Morrow, CC-BY 4.0.
Figure \(\PageIndex{3}\): These giant sequoia leaves were retrieved from a tree planted in a coastal environment, so the differences here represent evolutionary history, alone. Much like the coast redwood, these shiny leaves have a thick cuticle. The big difference to note is the thickness of these leaves, maintaining a much lower surface area to volume ratio than the coast redwood. The second thing to note is the orientation of the leaves. They are held much more closely together, reducing exposure to the wider environment. Two stomata are indicated in the image, though there are many. They are present on both adaxial and abaxial leaf surfaces. Photo by Maria Morrow, CC-BY 4.0.
There are also resin canals that ring the needle, appearing as holes surrounded by small cells. These secrete resin to protect the plant. Resin is a sticky fluid rich in compounds that protect the plant. It flows through canals in the stems, roots, and leaves and can rush to fill a wound. The resin can gum up the mouth parts of herbivorous insects, offer chemical defense against pathogenic bacteria and fungi, and harden to close the wound (much like a scab).
Reproduction in Conifers
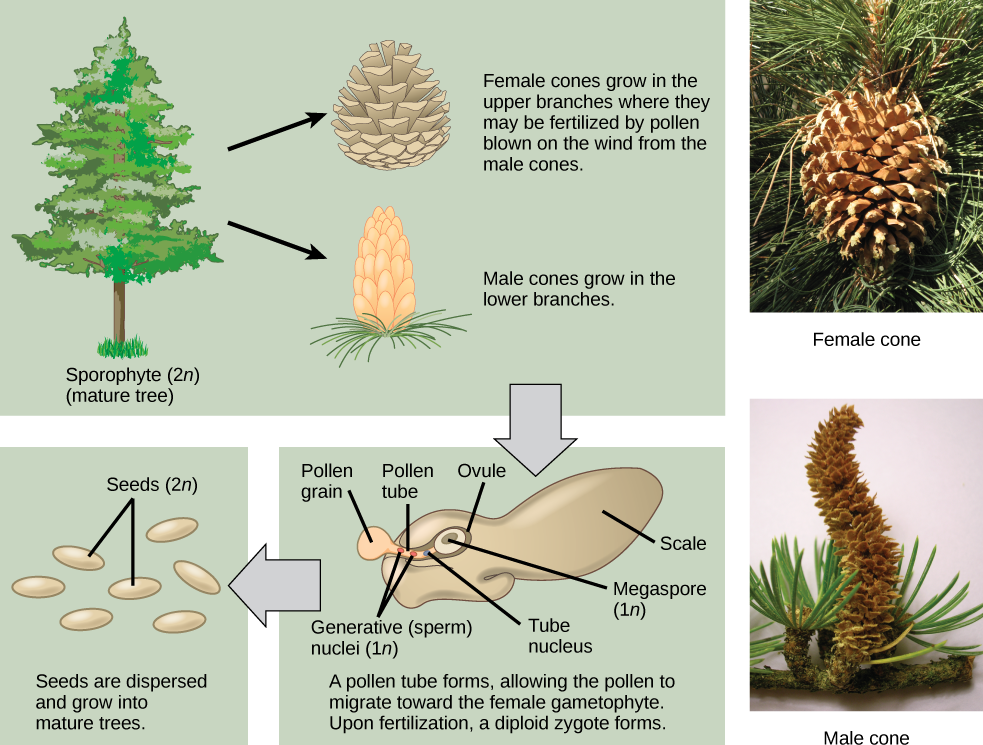
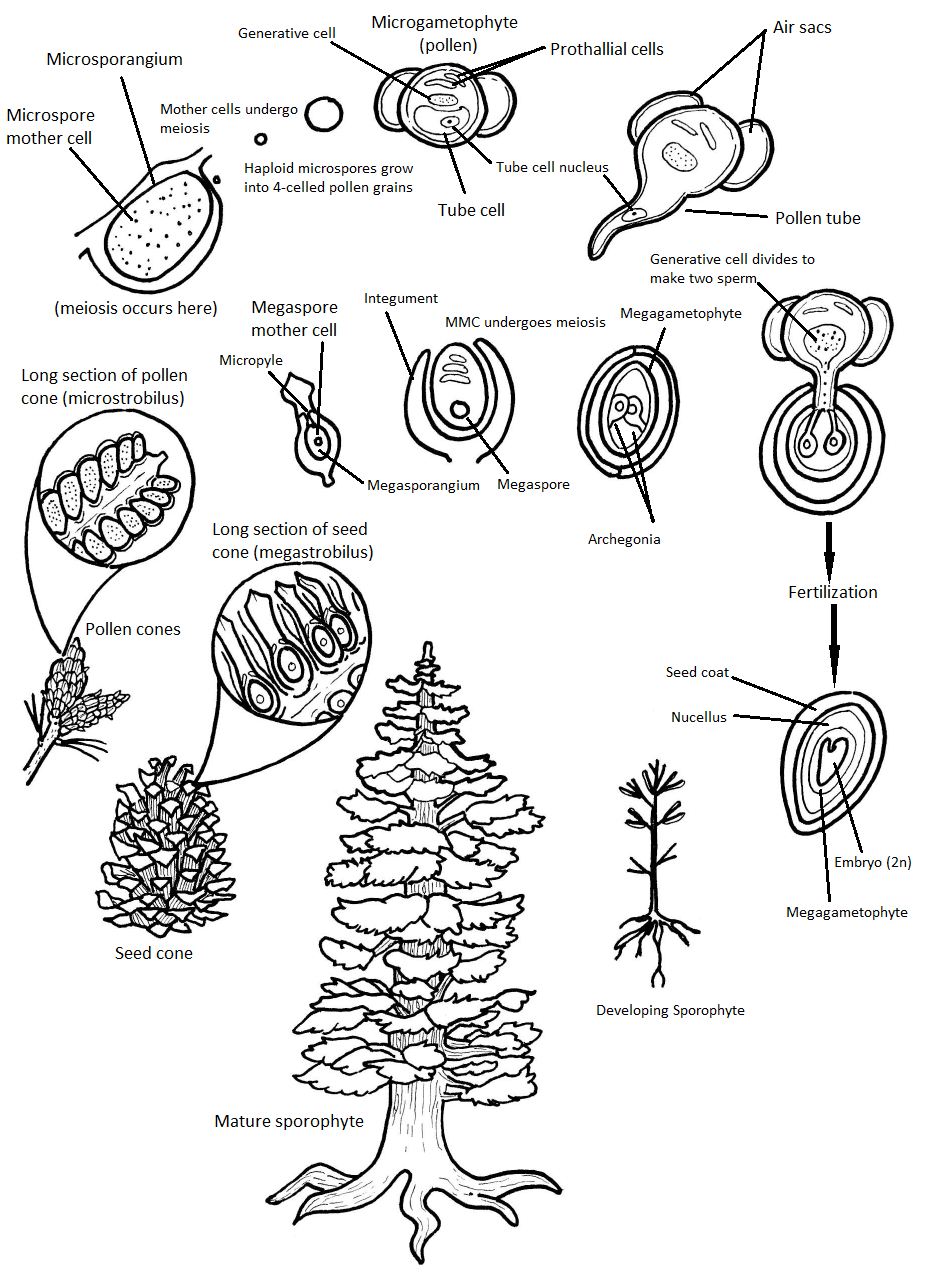
Unlike other gymnosperms, conifers are monoecious, meaning megastrobili and microstrobili are produced on the same plant. In general, megastrobili tend to be larger and longer-lived, while microstrobili are smaller and ephemeral, disintegrating after pollen is dispersed (see Figure \(\PageIndex{5-6}\)). Some conifers, like junipers (Juniperus) and yews (Taxus), lack woody cones and have fleshy scales. In all, conifer life cycle takes up to two years. Conifers do not have flagellate spermatozoa; their non-motile male gametes (spermatia) move inside long, fast-growing pollen tube.


The female cones are larger than the male cones and are positioned towards the top of the tree; the small, male cones are located in the lower region of the tree. Because the pollen is shed and blown by the wind, this arrangement makes it difficult for a gymnosperm to self-pollinate.
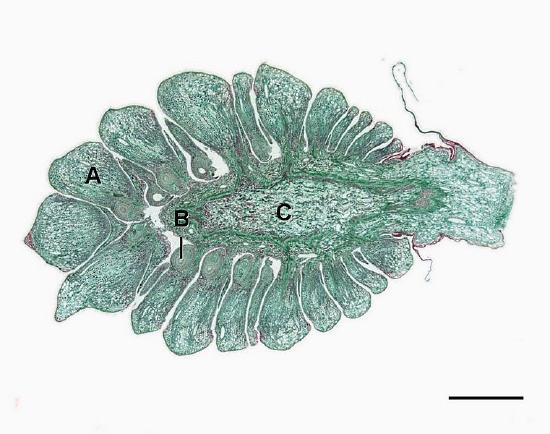
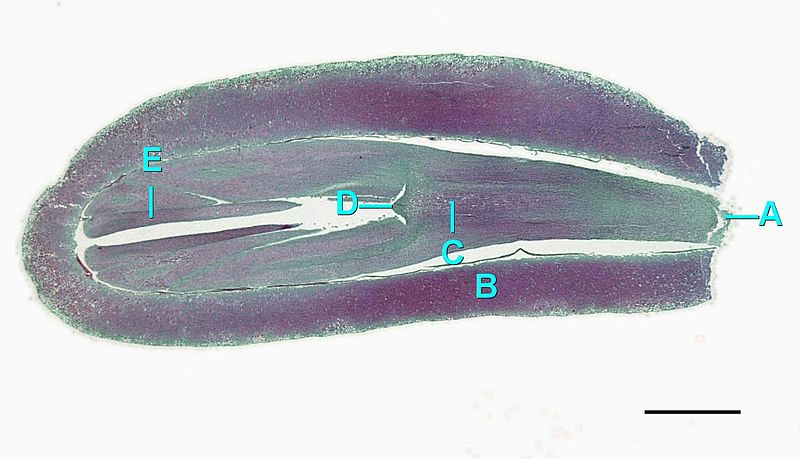
Seed Cones
The megastrobilus, or seed cone, is composed of spirally arranged megasporophylls called ovuliferous scales (Figure \(\PageIndex{7}\)). Each scale produces two megasporangia, which contain a diploid megasporocyte (also called a megaspore mother cell). Each megasporocyte undergoes meiosis. Only one of the four cells produced will survive to develop into a haploid megagametophyte and the other three will die. The megagametophyte is part of the ovule and contains archegonia, each with an egg cell inside (Figure \(\PageIndex{8}\)). The megagametophyte is retained within the megasporangium, which becomes the nucellus. Surrounding the nucellus is the integument, which is initially continuous with the ovuliferous scale and has a small opening called a micropyle.
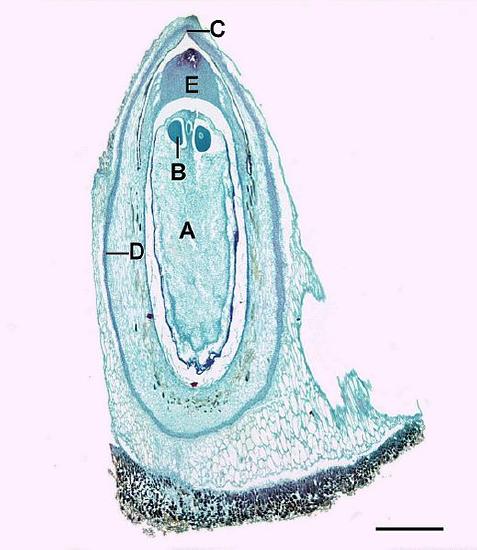
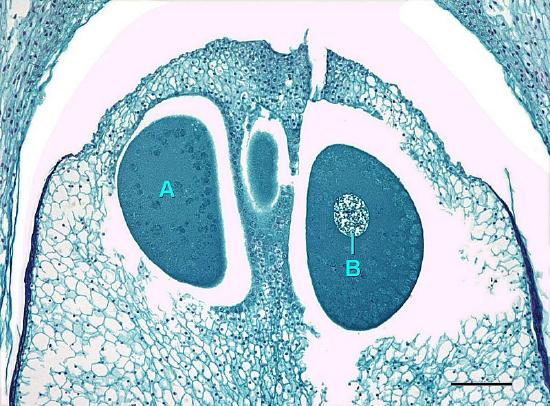
Pollen Cones
Microstrobili (pollen cones) are formed from overlapping microsporophylls that bear multiple microsporangia. Within the microsporangium, there are microsporocytes (also called microspore mother cells), diploid cells that undergo meiosis to produce haploid microspores. Microspores grow by mitosis into microgametophytes, AKA pollen, within the microsporangium. The microgametophyte in gymnosperms is the four-celled, "winged" pollen grain.
Within the pollen grain (Figure \(\PageIndex{9}\)), there is a generative cell, a tube cell, and two prothallial cells. On either side of the pollen grain, two wing-like structures called air sacs may help orient the pollen grain toward the ovule.
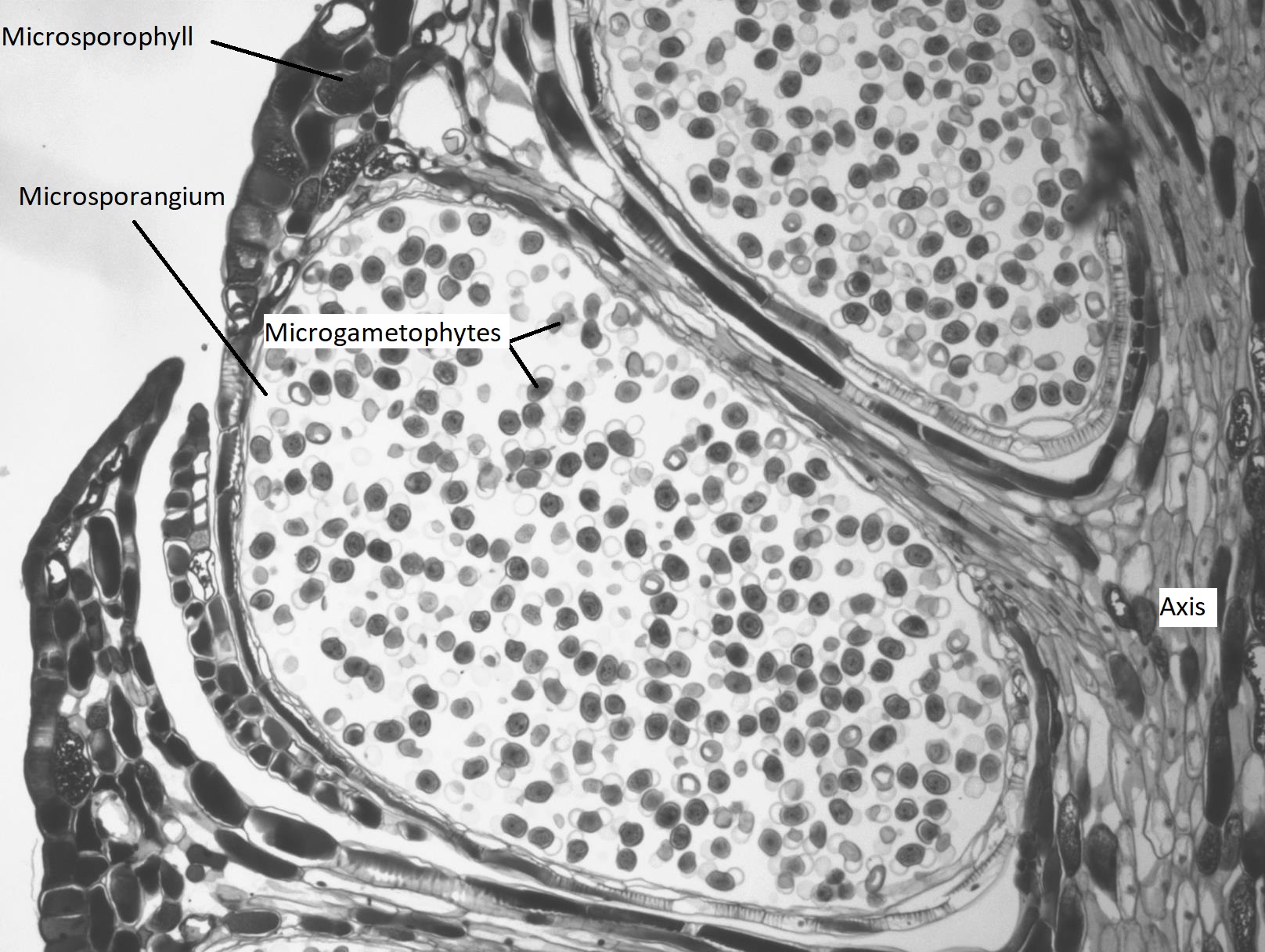
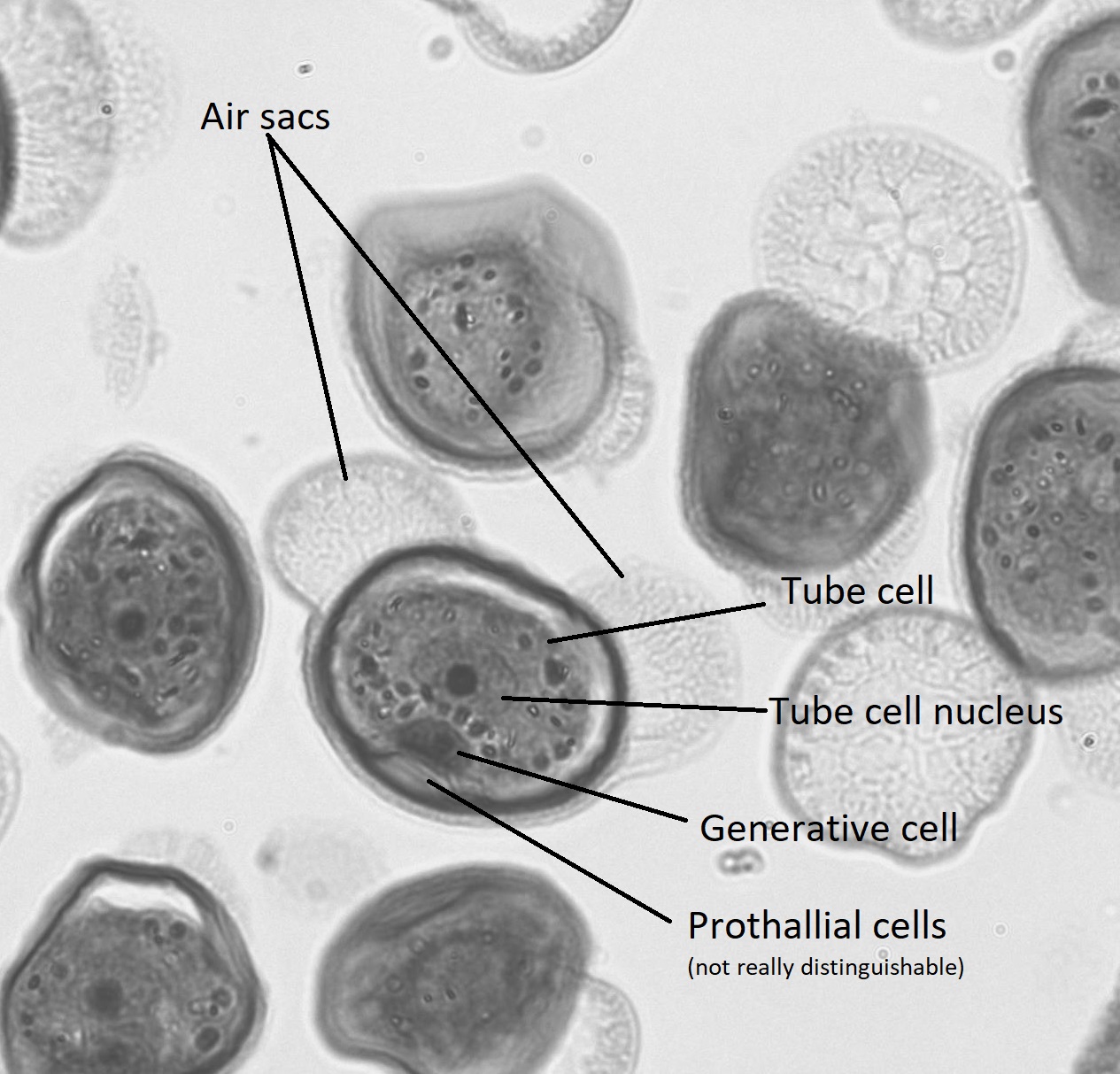
A grain of pollen will be transported on the wind and, if lucky, it will land on a seed cone. The seed cone has a drop of sugary liquid (a pollen drop) that it secretes, then retracts, pulling the pollen in toward the ovule. This stimulates the tube cell to germinate a pollen tube, while the generative cell divides by mitosis to produce two spermatia (no flagella). These spermatia travel with the pollen tube, through the micropyle, and into an archegonium where one will fertilize an egg (Figure \(\PageIndex{10-11}\)). It takes approximately one year for the pollen tube to grow and migrate towards the female gametophyte! When fertilization occurs, the micropyle closes and the integument becomes the seed coat.
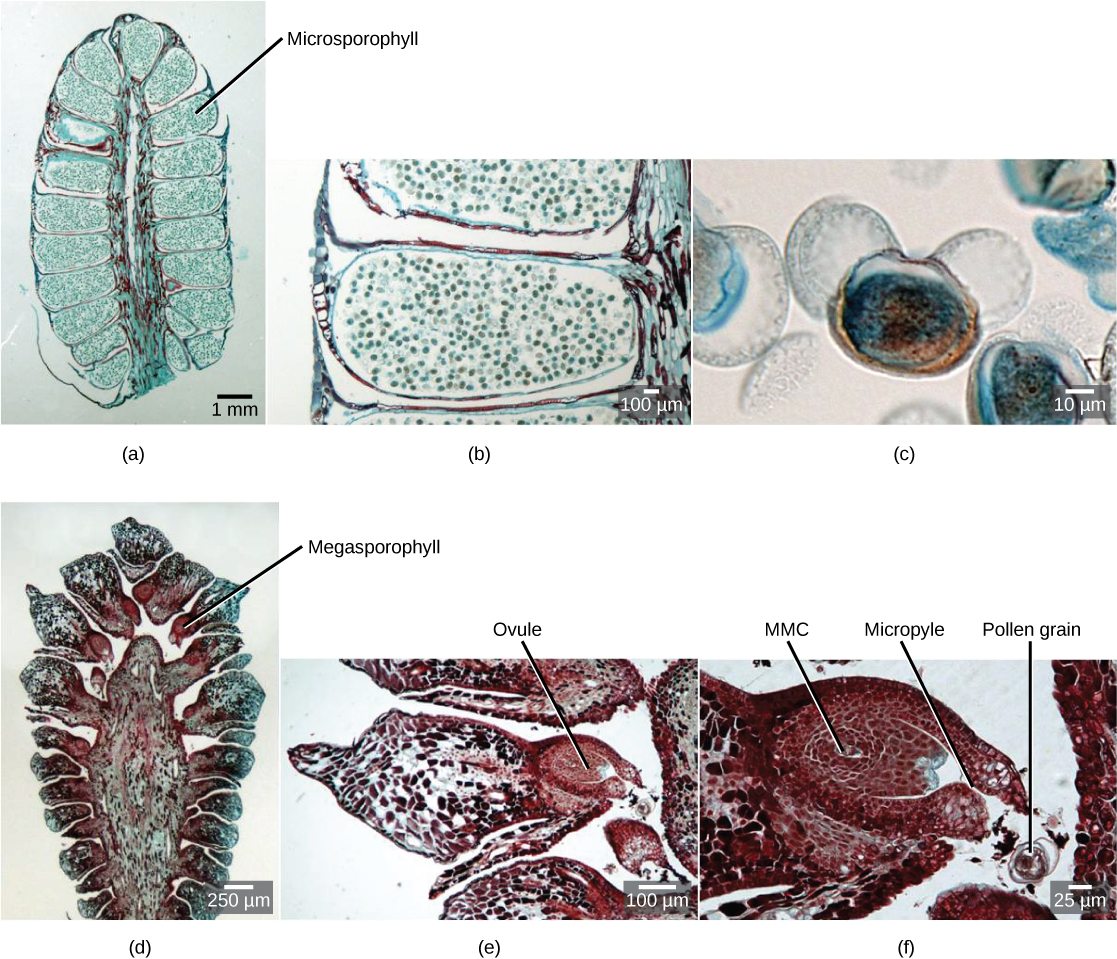

The zygote will grow and develop as an embryo within the seed, nourished by the megagametophyte tissue, as well as the nucellus (Figure \(\PageIndex{12}\)). The scales of the cones are closed during development of the seed. Seed development takes another one to two years. Once the seed is ready to be dispersed, the bracts of the female cones open to allow the dispersal of seed; no fruit formation takes place because gymnosperm seeds have no covering. The seed will be dispersed by wind or animals and germinate to grow into a diploid pine tree once again.
The Full Life Cycle
Video \(\PageIndex{1}\) is an extremely helpful narrated animation of the pine life cycle. You should watch this video or some other walkthrough of the pine life cycle before attempting to interpret the complex diagram (Figure \(\PageIndex{13}\)).
Video \(\PageIndex{1}\): A narrated video of the pine life cycle, sourced from Youtube.
Gnetophytes
Gnetophytes are a small group with only three genera that, excepting from their opposite leaves, seem not at all similar: Ephedra, Welwitschia, and Gnetum. Ephedra are horsetail-like desert leafless shrubs, Gnetum are tropical trees, and Welwitschia are strange desert plants that form two large, continuously growing leaves (Figure \(\PageIndex{14}\)). Gnetophytes represent an anatomically and genetically difficult group to classify. They have several traits in common with angiosperms, such as vessel elements in the xylem, double fertilization, and a covering over their seeds. Even their leaves are angiosperm-like, with netted venation. However, these traits are convergently evolved, meaning that angiosperms and gnetophytes each evolved these traits separately. Genetically, recent studies have placed the gnetophytes as a sister group to the Pinaceae (pine family) within the conifers. This would mean that pines, firs, and spruces are more closely related to strange gnetophytes like Ephedra than they are to other conifers like redwoods, cedars, and Pacific yew. However, the true nature of this evolutionary relationship remains murky and contentious.

Ephedra has archegonia, but in Gnetum and Welwitschia they are reduced. On the other hand, Ephedra and Gnetum have double fertilization, a process that you will see in angiosperms where both male nuclei fuse with cells of the one female gametophyte. Double fertilization in gnetophytes results in two competing embryos, and only one of them will survive in the future seed.
Both Gnetum and Welwitschia have vessel elements (like angiosperms). Gnetum also has angiosperm-like opposite leaves with netted venation, like the coffee tree (however, this probably is a result of modification of dichotomous venation). Ovules are solitary and covered with an additional outer integument; the male gametes are spermatia moving with the pollen tube instead of swimming (no flagella).
Welwitschia is probably most outstanding among gnetophytes. There is only one species and it occurs only in the Namibian desert. The best way to describe this plant is an “overgrown seedling.” It has a small trunk with two wide leaves that have parallel venation. The secondary thickening is anomalous, the wood has vessels. This plant is insect-pollinated and its winged seeds are dispersed by the wind. Fertilization is not double, but, along with pollen tubes, involves some interesting structures: prothallial tubes which grow from female gametophyte and meet with pollen tubes.
Characteristics of Gnetophytes
- Angiosperm-like features: vessel elements, double fertilization, fruit-like ovule coverings
- Dioecious. Female plants have covered ovules, while male plants have pollen cones.
- Leaves xerophytic with opposite arrangement
- Primarily insect pollinated; brightly colored seeds are dispersed by birds
Welwitschia mirabilis
This strange plant grows in the desert of Namibia. It has two large leaves that grow from a basal meristem. As the plant gets older the leaves split and start to look like numerous long tentacles (Figure \(\PageIndex{15}\)). The tips of the leaves are ragged, as these are the oldest parts. The leaves are shiny and the setting is dry, indicating their xerophytic nature. In the center, where the two leaves meet, plants will produce either megastrobili or microstrobili (Figure \(\PageIndex{16}\)).



Ephedra
Ephedra spp. have scale-like, opposite leaves produced on tough, photosynthetic stems. They produce swollen megastrobili that look like fruits (Figure \(\PageIndex{17}\)), and microstrobili have extruded microsporangia, making them look like catkins (a type of inflorescence produced by some angiosperms, (Figure \(\PageIndex{18}\)). Some Ephedra species produce alkaloids that have been extracted for stimulant use, including ephedrine and pseudoephedrine.
.jpg?revision=1&size=bestfit&width=758&height=568)

Gnetum
Gnetum spp. could easily be mistaken for flowering plants. They produce leaves with netted venation and fruit-like megastrobili. These plants are restricted to tropical areas and generally take on a tree-like habit.


Attribution
Curated and authored by Maria Morrow using the following sources:
- 19.1.5 Diversity and Evolutionary Relationships of the Plants from Biology by John. W. Kimball (licensed CC-BY)
- 26.2 Gymnosperms from Biology 2e by OpenStax (licensed CC-BY). Access for free at openstax.org.
- 7.6 Spermatophyta – Seed Plants from Introduction to Botany by Alexey Shipunov (public domain)