
2.3.2: Characteristics of Fungi
- Page ID
- 31934

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Learning Objectives
- List the characteristics of fungi.
- Describe the composition of the mycelium.
- Describe the mode of nutrition of fungi.
- Explain sexual and asexual reproduction in fungi.
The Fungal Body
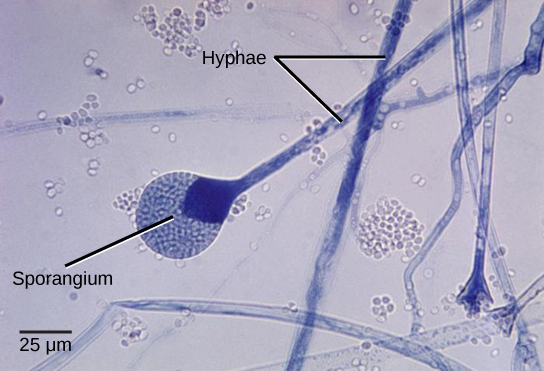
Fungi have well-defined characteristics that set them apart from other organisms. Most multicellular fungal bodies are made up of filaments called hyphae. Hyphae can form a network called a mycelium, which is the thallus (body) of the fungus (Figure \(\PageIndex{1}\)). Fungi are heterotrophs that excrete enzymes to digest food externally, then absorb the digested food. Because of this feeding strategy, fungi tend to live within whatever their food substrate is. The thin, filamentous hyphal structure allows for maximal surface area and thus, maximal contact with the food substrate.
.jpg?revision=1&size=bestfit&width=635&height=388)
Hyphae that have walls (septa) between the cells are called septate hyphae; hyphae that lack walls and cell membranes between the cells are called nonseptate or coenocytic hyphae (Figure \(\PageIndex{2-3}\)). Early diverging lineages of fungi have coenocytic hyphae, while later lineages (often referred to as "higher fungi") have septate hyphae.
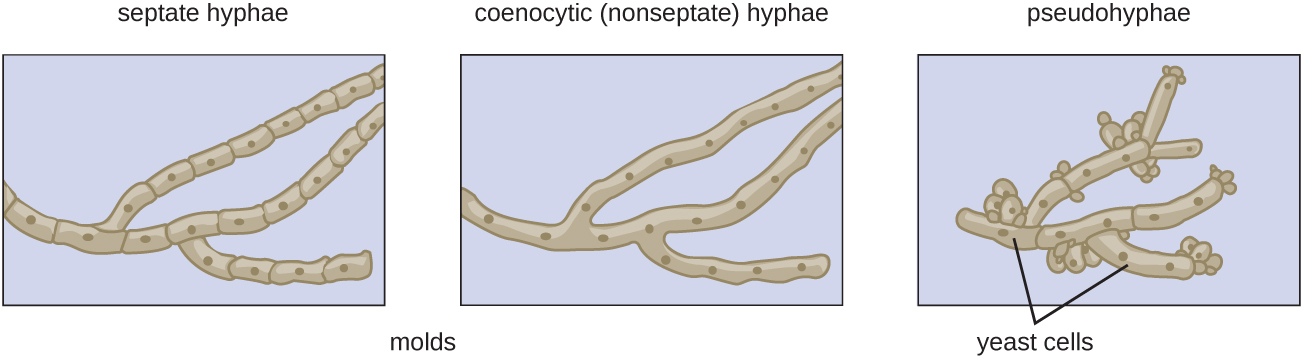

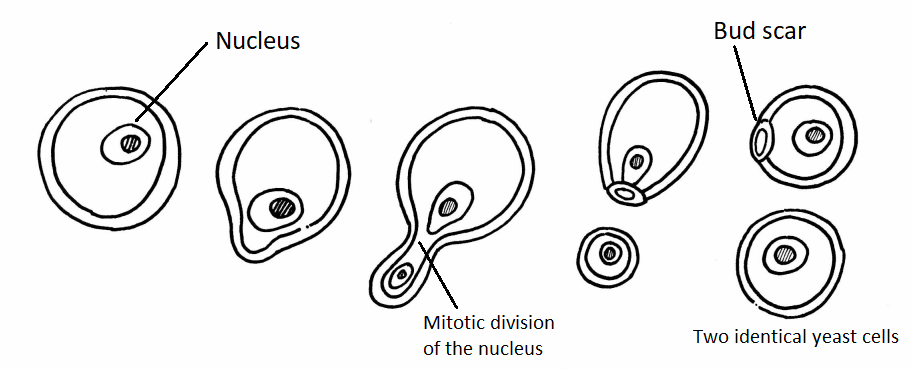
In contrast to filamentous fungi, yeasts are unicellular. The budding yeasts reproduce asexually by budding off a smaller daughter cell (Figure \(\PageIndex{4}\)); the resulting cells may sometimes stick together as a short chain or pseudohypha (Figure \(\PageIndex{2}\)). Candida albicans (Figure \(\PageIndex{5}\)) is a common yeast that forms pseudohyphae; it is associated with various infections in humans, including vaginal yeast infections, oral thrush, and candidiasis of the skin.

Some fungi are dimorphic, having more than one appearance during their life cycle. These dimorphic fungi may be able to appear as yeasts or filamentous forms, which can be important for infectivity. They are capable of changing their appearance in response to environmental changes such as nutrient availability or fluctuations in temperature, growing as a mold, for example, at 25 °C (77 °F), and as yeast cells at 37 °C (98.6 °F). This ability helps dimorphic fungi to survive in diverse environments.
Cell Structure and Function
Fungi are eukaryotes, and as such, have a complex cellular organization. As eukaryotes, fungal cells contain a membrane-bound nucleus. The DNA in the nucleus is wrapped around histone proteins, as is observed in other eukaryotic cells. A few types of fungi have structures comparable to bacterial plasmids (loops of DNA); however, the horizontal transfer of genetic information from one mature bacterium to another rarely occurs in fungi. Fungal cells also contain mitochondria and a complex system of internal membranes, including the endoplasmic reticulum and Golgi apparatus.
Unlike plant cells, fungal cells do not have chloroplasts or chlorophyll. Many fungi display bright colors arising from other cellular pigments, ranging from red to green to black, though none of these are for photosynthesis. The poisonous Amanita muscaria (fly agaric) is recognizable by its bright red cap with white patches (Figure \(\PageIndex{6}\)). Pigments in fungi are associated with the cell wall and some play a protective role against ultraviolet radiation. Some fungal pigments are toxic.

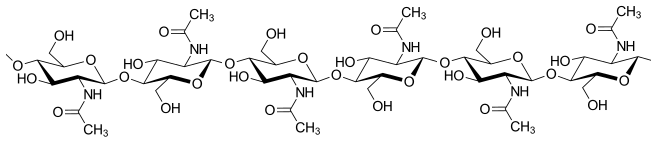
Like plant cells, fungal cells have a thick cell wall. The rigid layers of fungal cell walls contain complex polysaccharides called chitin and glucans (unlike the cell walls of plants, which contain cellulose). Chitin, also found in the exoskeleton of insects, gives structural strength to the cell walls of fungi. The wall protects the cell from desiccation and predators. Fungi have plasma membranes similar to other eukaryotes, except that the structure is stabilized by ergosterol, a steroid molecule that replaces the cholesterol found in animal cell membranes. Ergosterols are often exploited as targets for antifungal drugs.
Most members of the kingdom Fungi are nonmotile. Flagella are produced only in the chytrids. Motile cells have a single, whiplash flagellum, placing these organisms in the Opisthokonts (along with animals).
Growth
For filamentous fungi, a fungal spore germinates and grows apically. The mycelium is often highly branched and the fungus grows from the apex of each hypha, forming a radial growth pattern (Figure \(\PageIndex{8}\)). Because fungi grow within their substrate, we often do not see the mycelium, only the fruiting structures. In Figure \(\PageIndex{9}\), a circle of mushrooms called a fairy ring has formed in a grassy lawn. The original spore would have been deposited in the center of that ring and the mycelium grew outward from there. The mushrooms are produced at the edge of the mycelium.


Nutrition
Like animals, fungi are heterotrophs; they use complex organic compounds as a source of carbon, rather than fix carbon dioxide from the atmosphere as plants do. In addition, fungi do not fix nitrogen from the atmosphere. Like animals, they must obtain it from their diet. However, unlike most animals, which ingest food and then digest it internally in specialized organs, fungi perform these steps in the reverse order; digestion precedes ingestion. First, exoenzymes are transported out of the hyphae, where they process nutrients in the environment. Then, the smaller molecules produced by this external digestion are absorbed through the large surface area of the mycelium. As with animal cells, the polysaccharide of storage is glycogen, rather than starch, as found in plants.
Fungi are mostly saprotrophs, organisms that derive nutrients from decaying organic matter. They obtain their nutrients from dead or decomposing organic matter, mainly plant material. Fungal exoenzymes are able to break down insoluble polysaccharides, such as the cellulose and lignin of dead wood, into readily absorbable glucose molecules. The carbon, nitrogen, and other elements are thus released into the environment. Because of their varied metabolic pathways, fungi fulfill important ecological roles and are being investigated as potential tools in bioremediation. For example, some species of fungi can be used to break down diesel oil and polycyclic aromatic hydrocarbons (PAHs) due to their chemical similarity to the lignin found in wood. Other species take up heavy metals, such as cadmium and lead.
Some fungi are parasitic, causing infections in other organisms. Many important plant parasites are rusts, fungi in the Basidiomycota with complex life cycles. In environments poor in nitrogen, some fungi resort to predation of nematodes (small non-segmented roundworms). Species of Arthrobotrys fungi have a number of mechanisms to trap nematodes. One mechanism involves constricting rings within the network of hyphae. The rings swell when they touch the nematode, gripping it in a tight hold. The fungus penetrates the tissue of the worm by extending specialized hyphae called haustoria. Many parasitic fungi possess haustoria, as these structures penetrate the tissues of the host, release digestive enzymes within the host's body, and absorb the digested nutrients.
Some fungi live in a mutually beneficial symbiotic relationship with another organism, often a photosynthetic organism such as a plant or alga. The association of fungus and plant root is called mycorrhizae. Over 90% of land plant species benefit from symbiotic mycorrhizal relationships. The plant benefits by more-efficient mineral (e.g. phosphorus) uptake and the fungus benefits by the sugars translocated to the root by the plant.
Mycorrhizal fungi may also form conduits for nutrients between plant species, for better or worse. The colorless, and hence heterotrophic ghost pipe (Monotropa uniflora - pictured in Figure \(\PageIndex{10}\)) is an angiosperm that must secure all its nourishment from mycorrhizal fungi that are attached at the same time to the roots of some autotrophic plant such as a pine tree. Radioactive carbon administered to the pine (as CO2) soon turns up in carbohydrates in nearby ghost pipes.

Reproduction
Fungi disperse themselves by releasing spores (haploid, in most cases). Fungal spores are present almost everywhere (and are a frequent cause of allergies). Spores of the wheat rust fungus have been found at 4000 m in the air and more than 1450 km (900 miles) from the place they were released. No wonder then that many fungi seem to have a worldwide distribution.

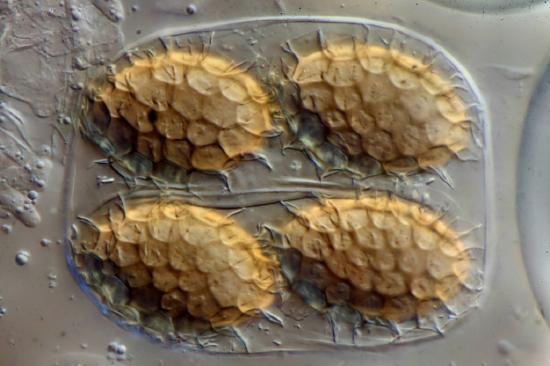
Fungi reproduce sexually and/or asexually. Fungi may exhibit asexual reproduction by mitosis with budding (Figure \(\PageIndex{13}\)), fragmentation of hyphae, and formation of asexual spores by mitosis (Figure \(\PageIndex{14}\)). Depending on the taxonomic group, sexually produced spores are known as zygospores (in the former Zygomycota), ascospores (in Ascomycota), or basidiospores (in Basidiomycota).
In both sexual and asexual reproduction, fungi produce spores that disperse from the parent organism by floating on the wind, hitching a ride on an animal, or some other means. Fungal spores are microscopic and often produced in large numbers. When the giant puffball mushroom bursts open, it releases trillions of spores. The huge number of spores released increases the likelihood of landing in an environment that will support growth (Figure \(\PageIndex{12}\)).

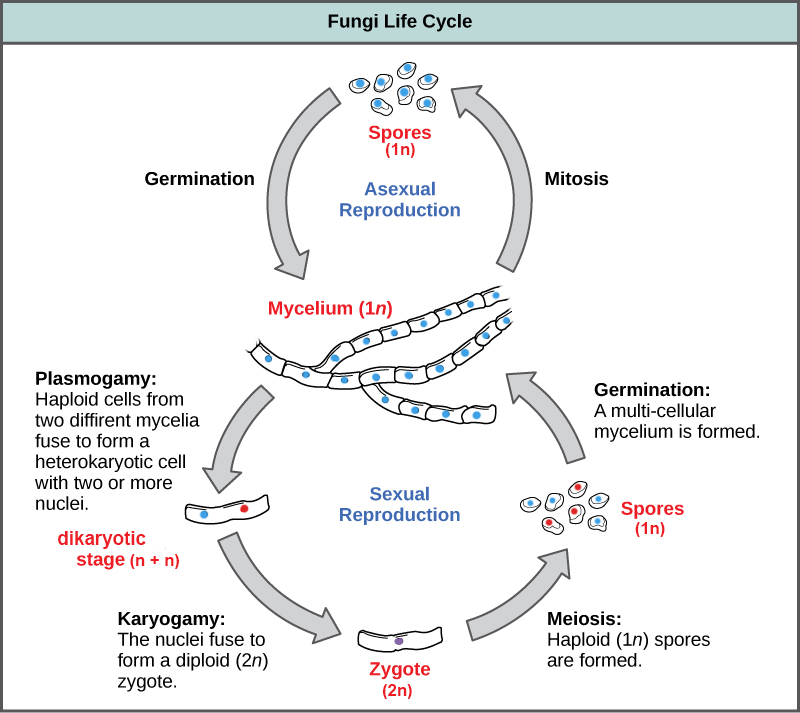
Fungal life cycles are unique and complex, though fungi are typically said to have a haplontic life cycle (Figure \(\PageIndex{15}\)). In a generalized fungal life cycle, haploid mycelia of different mating types either form hyphae that have gametes at the tips or fuse directly (somatogamy). The cytoplasm fuses in an event called plasmogamy, producing a cell with two distinct nuclei (a dikaryotic, or n+n, cell). At some point later, the nuclei fuse (in an event called karyogamy) to create a diploid zygote. The zygote undergoes meiosis to form haploid spores that germinate and grow into a new haploid mycelium. How these events occur is one of the major ways we classify fungi, and the life cycles of different fungal groups contrast significantly.

Review the characteristics of fungi by visiting this interactive site from Wisconsin-online.
Summary
Fungi are eukaryotic organisms that appeared on land more than 450 million years ago. They are heterotrophs and contain neither photosynthetic pigments such as chlorophyll, nor chloroplasts. Fungi are important decomposers that release essential elements into the environment. However, they have many other important roles in ecosystems as parasites and mutualists. External enzymes digest nutrients that are then absorbed by the mycelium of the fungus, which is composed of filamentous cells called hyphae. A thick cell wall made of chitin surrounds the hyphae. Unicellular fungi are called yeasts and reproduce by budding. Fungi can reproduce asexually (typically referred to as molds) and/or sexually. One of the primary ways fungi were classified into phyla was based on the life cycle and type of sexual structures produced.
Characteristics of Fungi:
- Eukaryotic
- Heterotrophic by absorption
- Morphology: Unicellular or a thallus composed of hyphae. If present, motile cells have only a single, whiplash flagellum.
- Cell walls: Chitin
- Storage carbohydrate: Glycogen
- Life cycle: Haplontic (though this one gets a bit weird)
- Ecology: Many fungi are decomposers, primarily in terrestrial ecosystems. Other groups of fungi live in symbiosis with other organisms as parasites, mutualists, or commensalists.
Attribution
Curated and authored by Maria Morrow, CC-BY-NC, using the following sources:
- 19.1.7 Fungi from Biology by John. W. Kimball (licensed CC-BY)
- 24.1 Characteristics of Fungi from Biology 2e by OpenStax (licensed CC-BY). Access for free at openstax.org.
- 5.3 Fungi from Microbiology by OpenStax (licensed CC-BY). Access for free at openstax.org.