
2.4.2.1: Slime Molds
- Page ID
- 37000

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Learning Objectives
- Differentiate between the three major groups of slime molds.
- Differentiate between slime molds, fungi, and plants.
- Identify structures and phases in the myxomycete life cycle and know their ploidy.
Slime molds are an unusual group of organisms that have previously been classified as animals, fungi, and plants. Like plants, slime molds have cellulose in the cell walls of their spores. Unlike plants, slime molds are heterotrophs! Though they were formally classified as fungi, slime molds do not have chitin in their cell walls and have a diplontic life cycle (Figure \(\PageIndex{12}\)). These organisms move about as amoebae engulfing bacteria (unlike fungi, who digest food externally). When conditions become unfavorable, whether due to lack of food or lack of moisture, they form spores. They can be found in damp substrates with ample bacteria and are most frequently found on decaying logs and forest duff.
There are several different lineages of organisms commonly referred to as slime molds. Cellular slime molds (dictyostelids, Figure \(\PageIndex{1}\)(a)) are groups of unicellular amoebae that collaborate to form fruiting structures to disperse spores. Protostelids make small fruiting bodies that have cellular stalks. Plasmodial slime molds (classified as Myxogastria or Myxomycetes, Figure \(\PageIndex{1}\)(b)) form a large, multinucleate amoeba with no cell wall that will eventually wall off individual nuclei to form spores.

Dictyostelids
The cellular slime molds exist as individual amoeboid cells that periodically aggregate. The individual amoebe can be seen aggregating in Figure \(\PageIndex{1}\)(a). The aggregate then forms a fruiting body (Figure \(\PageIndex{2}\)) that produces haploid spores. One cellular slime mold, Dictyostelium discoideum, has been an important study organism for understanding cell differentiation, because it has both single-celled and multicelled life stages, with the cells showing some degree of differentiation in the multicelled form. Watch Video \(\PageIndex{1}\) to see how these individuals aggregate into a single fruiting body.

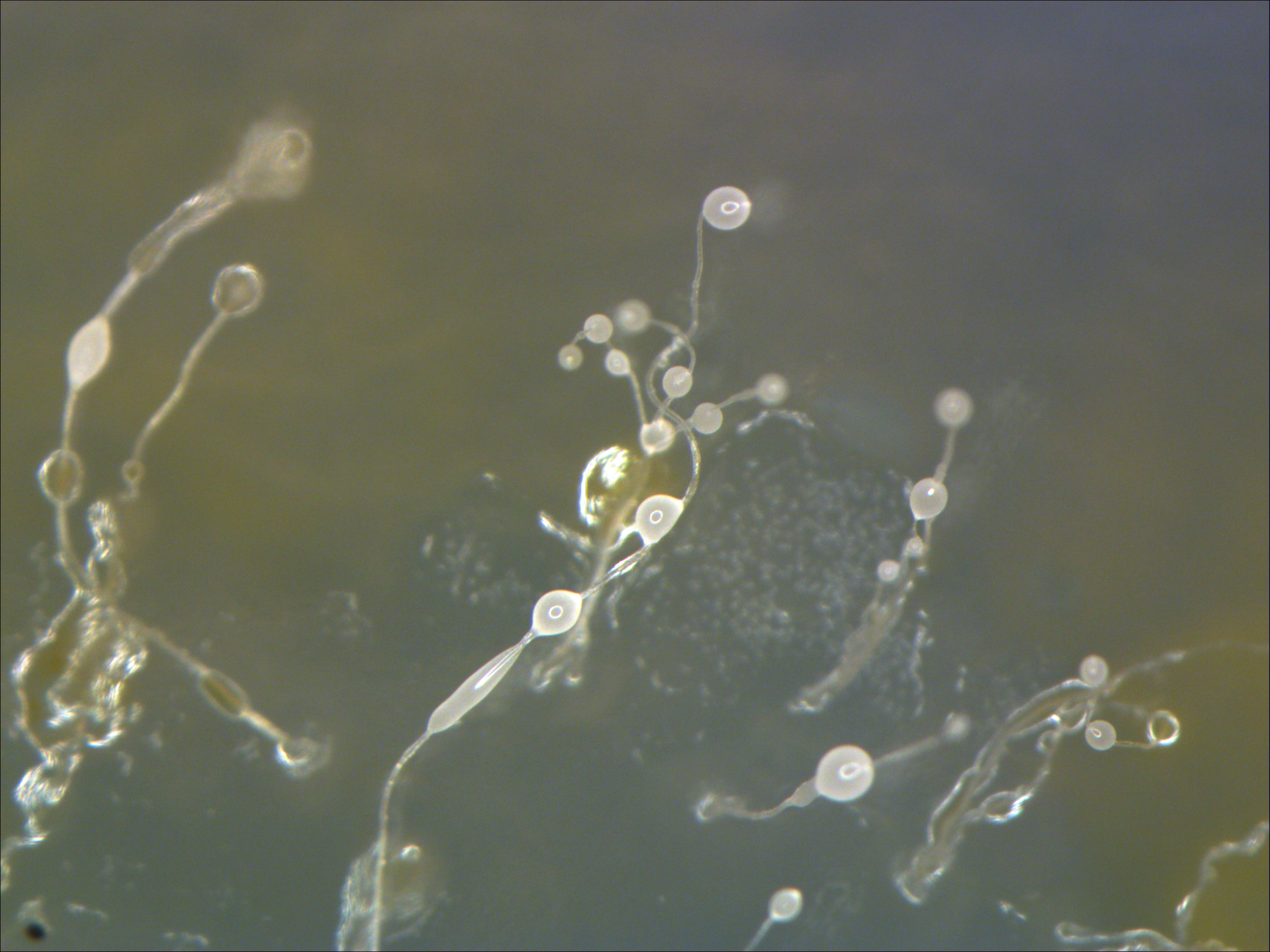
Video \(\PageIndex{1}\): Watch the strange behavior of the cellular slime mold Dictostelium discoideum as individual amoebae respond to an aggregation signal (cAMP), form a mobile slug, and eventually produce a stalked fruiting structure and spores. Sourced from YouTube.
The organisms in this group have a complex life cycle (Figure \(\PageIndex{3}\)) during the course of which they go through unicellular, multicellular, spore producing, and amoeboid stages. Thousands of individual amoebae aggregate into a slimy mass - each cell retaining its identity (unlike plasmodial slime molds). The aggregating cells are attracted to each other by the cyclic AMP (cAMP) that they release when conditions become stressful, such as a depletion in food. Individual amoebae respond to the chemical signal by moving to areas of higher cAMP concentration (chemotaxis), eventually aggregating into a single slug. The slug can respond to moisture and light gradients, navigating to a good spot for spore production. Some cells in the slug contribute to a 2–3-millimeter stalk, drying up and dying in the process. Cells atop the stalk form an asexual fruiting body that contains haploid spores. The spores are disseminated and can germinate if they land in a moist environment.
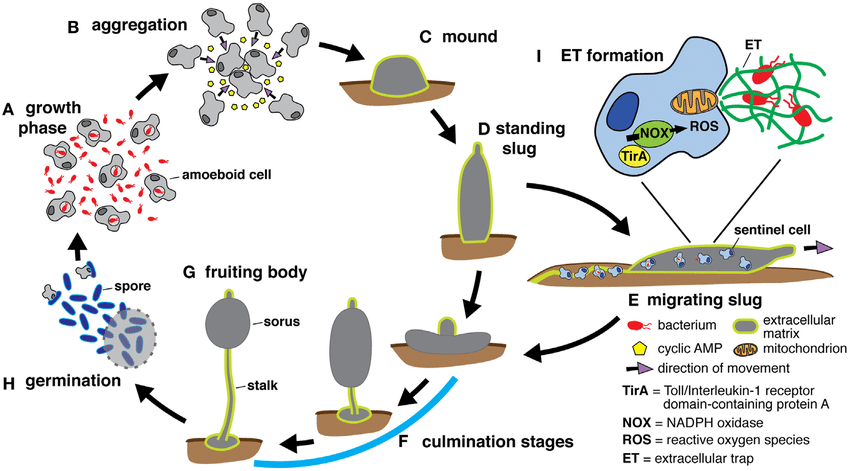
Protostelids
Protostelids are a group that has received less attention than either the Dictyostelids or plasmodial slime molds, as each of the latter groups contains a model organism used to study a specific system. Protostelids make simple fruiting bodies, similar to the Dictyostelids, with a stalk and spores at the apex. The slime mold Ceratiomyxa looks more like a plasmodial slime mold, but closer inspection reveals that spores are formed on minute, stalked fruiting bodies covering the external surface of the tentacle-like structures (Figure \(\PageIndex{5}\)). Ceratiomyxa may not actually be a protostelid, but the small, stalked fruiting bodies formed on the external surface are similar to what would be found in a true protostelid.


Plasmodial Slime Molds (Myxogastria)
Plasmodial slime molds represent a vast diversity of morphologies. While still a plasmodium (see Figure \(\PageIndex{6}\)), they can be difficult to distinguish. However, once they have formed into a fruiting structure, they can form distinct, varied, and amazing shapes (see Figures \(\PageIndex{7-10}\))!
The Plasmodium
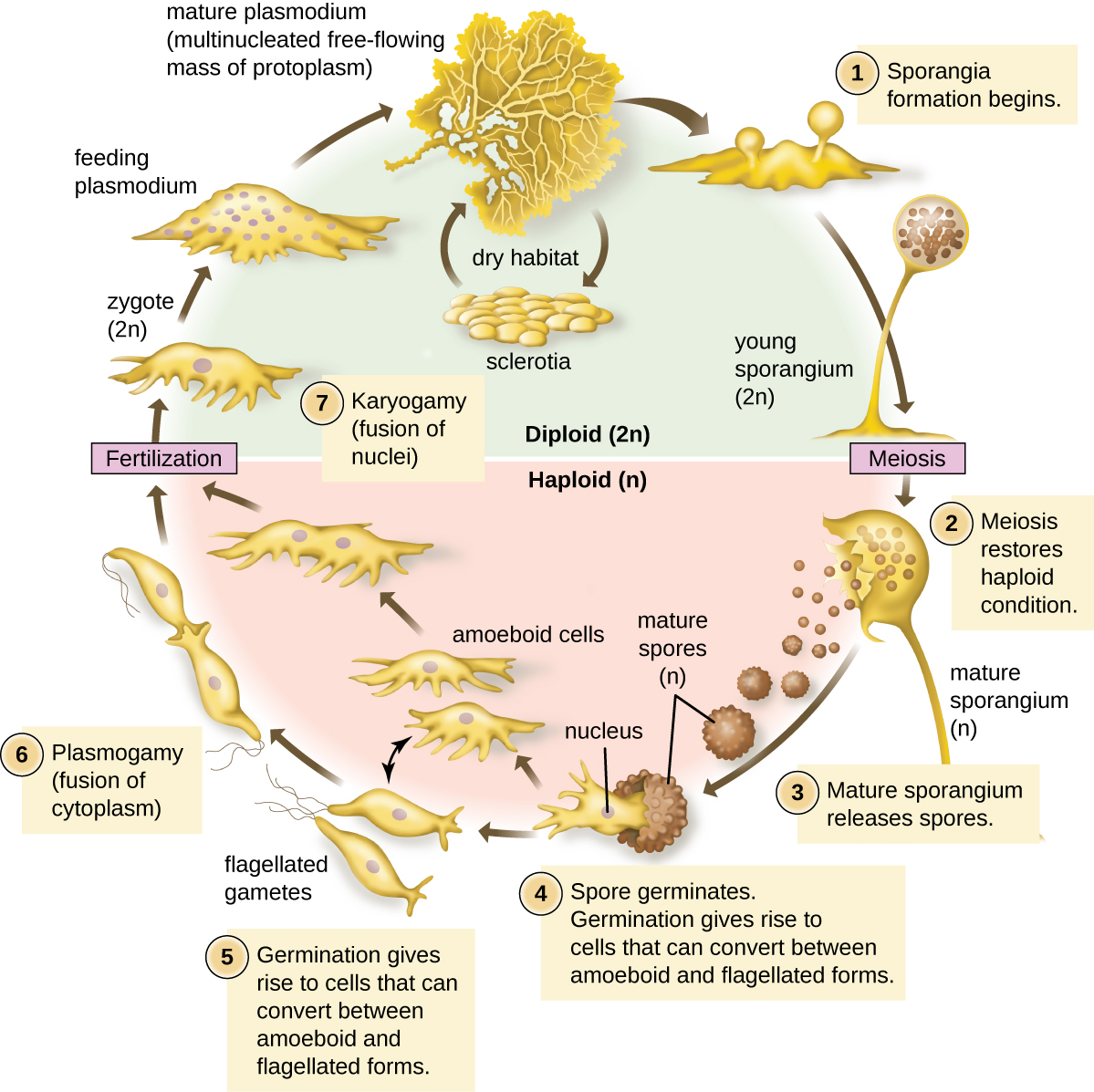
In their feeding stage, myxomycetes form one large amoeba called a plasmodium with many nuclei and no cell wall. This plasmodium moves over damp, decaying material looking for bacteria (and sometimes fungi) to engulf and digest. When it dries out or runs out of food, it begins to make fruiting structures called sporangia (sporangium, singular). Inside these sporangia, the diploid nuclei will undergo meiosis and haploid nuclei will be walled off to make spores for aerial dispersal. Dispersal by spores, heterotrophism, and glycogen as a storage carbohydrate originally classified this group within Kingdom Fungi, but this is the end of the similarities. The spores have cell walls made of cellulose, like plants. When these spores land, they will germinate into haploid cells with two flagella (called swarm cells) or amoebae that will fuse together to form a diploid plasmodium. See Figure \(\PageIndex{11}\) for a diagram of this life cycle.

Sporocarp Diversity
The diversity of sporulating structures, or sporocarps, has led many to fall in love with this group of organisms. In Hemitrichia serpula, the plasmodium forms into a network of veins that then become fruiting structures (a plasmodiocarp, see Figure \(\PageIndex{7}\)). In some slime molds, like Fuligo and Lycogala, the entire plasmodium forms a cushion that dries and produces spores (an aethalium, see Figure \(\PageIndex{8}\)). In other slime molds, individual sporangia are so closely clustered together, they appear to be a single fruiting structure (a pseudoaethalium, see Figure \(\PageIndex{9}\)). The last type of sporocarp is more familiar, forming many distinct stalked sporangia (see Figure \(\PageIndex{10}\)).




Life Cycle
The life cycle of plasmodial slime molds is best classified as diplontic: the "multicellular" (actually just multinucleate) phase is diploid. Haploid cells that germinate from spores (amoebae or biflagellate swarm cells) do not grow until after they have fused with another haploid cell. In some myxomycetes, amoebae or swarm cells produced from the same parent plasmodium can fuse together to form a new plasmodium. This is called homothallism (homo- meaning same, thallus). In other myxomycetes, these gametes must be from different individuals (heterothallism, hetero- meaning other). The discovery of different mating types in myxomycetes, as well as the genes that determine mating type, was made by O'Neil Ray Collins (Figure \(\PageIndex{11}\)).

Summary
Slime molds represent several different lineages: the cellular slime molds (Dictyostelids), Protostelids, and plasmodial slime molds (Myxomycetes). These organisms move about as amoebae consuming bacteria until conditions become unfavorable, at which point they form spores. They can be found in damp substrates with ample bacteria and are most frequently found on decaying logs and forest duff.
Dictyostelids are model organisms for studying altruism. They are unicellular, but collaborate to form multicellular structures where only some of the individuals involved go on to make spores. Protostelids are less well-understood and form a single sporangium at the tip of a cellular stalk. Plasmodial slime molds (the myxomycetes) form a large, multinucleate amoeba during their feeding stage called a plasmodium. They have diplontic life cycles and there is a lot of morphological diversity of sporocarps represented in this group. Some organisms in this group are studied for their ability to solve mazes and spatial puzzles.
Though these organisms seem primitive, they have complex interactions with each other and their environments.
Attributions
Curated and authored by Maria Morrow, CC BY-NC, using the following sources:
- 19.1.2 Protists from Biology by John. W. Kimball (licensed CC-BY)
- 23.3 Groups of Protists from Biology 2e by OpenStax (licensed CC-BY). Access for free at openstax.org.
- 5.1 Unicellular Eukaryotic Organisms from Microbiology by OpenStax (licensed CC-BY). Access for free at openstax.org.
Tags recommended by the template: article:topic