
11.1: Membrane and Membrane Proteins
- Page ID
- 14983


Introduction
One easily understandable function of membrane bilayers is to separate the inside and outside of the cell or intracellular organelles. Yet as we mentioned before, such barriers can not be so rigid and impenetrable that they prevent the movement of materials across the membrane. Also, all cells must sense and respond to their environment through a process called signal transduction. We have already discussed lipid molecules involved in signaling. Now let's turn our attention to proteins that associate with the membrane and confer added functionalities to it. Figure \(\PageIndex{1}\) reviews some of the features of membranes we've discussed before and shows a simple bilayer (top) to the complicated membrane/cell wall of bacteria.

Types of Membrane Proteins
Although we presented this image earlier, Figure \(\PageIndex{2}\) reviews the details that should now be clearer to you. In this section, we will explore membrane proteins in more detail.

Proteins can be loosely associated with the membrane (peripheral or extrinsic) or can embed deeply and most typically pass through the membrane and become a transmembrane (also called integral or intrinsic) protein. Sometimes they pass through using a single alpha helix, while other times they pass through multiple times (for example seven times in G-protein coupled receptors). They can also be classified based on the number of leaflets of the membrane they cross, as shown in Figure \(\PageIndex{3}\).

Peripheral Proteins
The proteins interact with a membrane through protein-lipid head group interactions, but might slightly penetrate the membrane. Those that do would be classified as monotopic peripheral. Peripheral proteins are generally easy to remove from a membrane in vitro by changing solution ions concentration as the interactions are often ion-ion nature. The first model below shows the binding of a matrix Metalloproteinase (MPP) 12 to a lipid bilayer. This protein is involved in inflammation, wound healing, arthritis, cardiovascular disease, and remodeling of neural synapses, suggesting a broad role in recovery from cell and tissue aberrations. MMP-12 is secreted by macrophages so it is considered a water-soluble (aqueous) protein. It travels to viral cells and appears to display activity not in aqueous solution but near membranes, implying activation of the enzyme through binding to the bilayer.
Studies show the catalytic domain of MMP 12 can bind bilayers through both α- and β- secondary structure regions of the protein. Figure \(\PageIndex{4}\) shows an interactive iCn3D model of the protein and its interaction with the membrane through the alpha-helical region. Once bound to the membranes, catalytic activity increases.
_12%25C2%25A0_lipid_bilayer..png?revision=1)
Other MMPs localize to membranes in other ways. MMP-7 interacts with heparan sulfate proteoglycans (CD44) and lipid rafts. Others bind transmembrane proteins like integrins.
Other examples of peripheral protein include many precursor forms of protein clotting factors. Clotting is initiated when the serine protease thrombin cleaves fibrinogen to form fibrin, which self-associates to form a fibrin clot, or when thrombin activates receptors in platelets. The soluble precursor of thrombin, prothrombin, a zymogen, is activated on membrane binding through interactions with several proteins assembled on a negatively charged phospholipid (like phosphatidylserine) bilayer in the prothrombinase complex. How does the precursor zymogen interact with the membrane? It requires calcium ions, which bind to a series of gamma-carboxylated glutamic acid (GLA) residues on the zymogen. The enzyme that carboxylates the zymogen depends on Vitamin K.
Figure \(\PageIndex{5}\) shows an interactive iCn3D model of bovine prothrombin Fragment 1 (N terminal) bound to a bilayer through its GLA domain (1NL2).
.png?revision=1&size=bestfit&width=188&height=318)
The Gla sidechains are shown in CPK-colored sticks and interact with Ca2+ ions (gray spheres). Click on this link to see a zoomed view of just the calcium ions and Gla sidechains: https://structure.ncbi.nlm.nih.gov/i...xydtj8a4WHgdb7
The Gla domain in the absence of calcium ions is disordered. On binding, an ordered linear alignment of bound calcium ions is formed, stabilizing the ordered structure of the Gla domain and allowing interaction with the membrane. Three nonpolar amino acid side chains, Phe 5, Leu 6, and Val 9, are now clustered and exposed, allowing penetration of this hydrophobic patch part-way into the membrane. They are represented in cyan spacefill just underneath the surface of the red dots in the model above (the red dots are dummy atoms that represent the outer bilayer leaflet). Given this penetration, this protein domain would then be considered monotopic.
What is not shown in the model is the role of negatively charged phosphatidyl serine. Studies have shown that the head group of serine in lysophosphatidylserine (which has only one acyl group) provides additional ion-ion interactions with the Ca2+ ions that also bind Gla residues 17 and 21. Arg 10 and Arg 16 also interact with the phosphatidyl serine head group. Phosphatidylcholine could also spatially fit into the active site but electrostatic interactions would prevent it. Why?
Lipid-Anchored Proteins
We have studied lipids, proteins, and carbohydrates. Although phospholipids can spontaneously form bilayers, the actual structure of biological membranes is made much more complicated through the addition of protein and carbohydrate substituents to the membrane. Soluble proteins can be made to insert into bilayers by the addition of nonpolar attachments. Localization to a membrane changes the functional expression of the protein. Several examples of such attachments are described below.
Fatty acid linkers
Two common covalent modifications of proteins are N-myristoylation (attached myristic acid - 14:0 - through an amide link) and S-palmitoylation (attached palmitic acid - 16:0 - through a thioester link with a Cys).
Myristoylation is usually a cotranslational modification in eukaryotic and viral proteins that occurs after cleavage of the N-terminal methionine. Figure \(\PageIndex{6}\) shows an image of the serine/threonine phosphatase 2C (1A6Q) with its N-terminal glycine myristoylated. It should be obvious how this post-translationally modified protein interacts with a membrane.
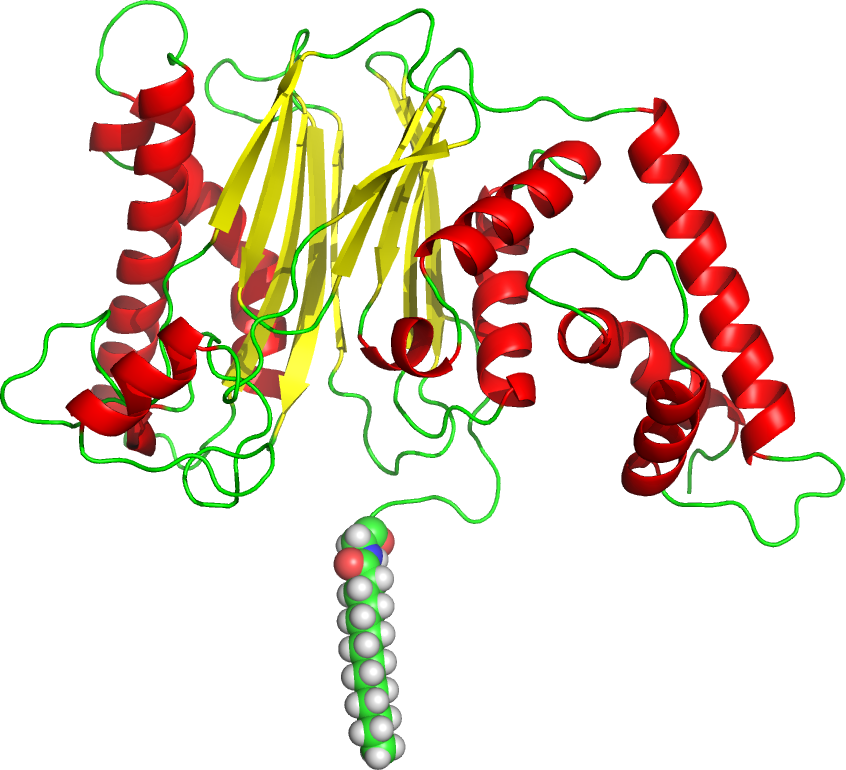
Figure \(\PageIndex{6}\): The serine/threonine phosphatase 2C (1A6Q) with its N-terminal glycine myrisoylated
This modification is a key part of initiating immune system signal transduction pathways. The modification is catalyzed by N-myristoyltransferase (NMT) using myristoyl-coenzyme as the fatty acid acyl donor. This activates the function of the protein in part by reducing the dimensionality of substrate diffusion to the protein to the 2D surface of the membrane instead of a 3D search in the cytoplasm. NMT acylates protein at this consensus sequence: G1X2X3X4S/T5X6R7R8.
Likewise, many signaling proteins are palmitoylated, leading to protein recruitment to membranes. Small G proteins like Ras, Rho, and the alpha subunit of heterotrimeric G proteins are often palmitoylated. This modification is also be found in transmembrane proteins in which localization is not an issue (see example xx below). In such circumstances, the modification might however help in targeting the proteins to rafts within the membrane. Palmitic acid is saturated and the addition of it to a protein might target it to more ordered regions of the membrane with cholesterol and sphingolipids within rafts.
Isoprenoids linkers:
The isoprenoids farnesyl (15C) or geranylgeranyl (20C)are added to a CAAX carboxy-terminal sequence in a target protein like RAS, where C is Cys, A is aliphatic, and X is any amino acid, which helps target proteins to the membrane. The enzymes used for these modifications are farnesyltransferase (FTase) and protein geranlygeranyltransferase
I (GGTase I), respectively. For this and the other modifications, it has the potential to do more than target proteins to the membrane. The modification can also modulate protein-ligand interactions and protein stability. Ras, a key signaling protein, is a target of prenylation.
Ras and other small G proteins are involved in a large percentage of human cancers. As the G protein Ras has somewhat of a billiard ball surface with obvious sites to target drugs that would affect its aberrant function in cancers, efforts have been made to target the prenyltransferases necessary to target it to the membrane.
In humans, there are 3 different genes in the Ras family, H-Ras and N-Ras, whose gene products localize to both plasma and Golgi membranes, and K-Ras, which localizes predominantly to the plasma membrane. These and other G proteins bind GTP and possess GTPase activity. The GTP-bound form is active, while the GDP form is inactive. Point mutations that attenuate or prevent GTP cleavage leave the protein continually activated which contributes to oncogenesis.
KRas has two predominant isoforms, 4A, the canonical form (also called 2A), and 4B (also called 2B) that arise from alternative splicing of the primary RNA transcript. The C-terminal protein sequences of isoform 4A and 4B differ significantly.
Isoform 4A: QYRLKKISKEEKTPGCVKIKKCIIM
Isoform 4B: KHKEKMSKDGKKKKKKSKTKCVIM
The farnesylation motif site containing the modified Cys are highlighted in yellow above. The same cysteine is also often carboxymethylated. The Cys six residues from the farnesylated Cys in isoform 4A are also often palmitoylated
Figure \(\PageIndex{7}\) shows isoform KRas 4B bound to a membrane bilayer through its farnesylated tail. (PDB file provided by Alemayehu (Alex) Gorfe. Viney Nair and Andrew McCammon). The tail is essential for its function at the plasma membrane where KRAS-mediated signaling events occur. Phosphodiesterase-δ (PDEδ) binds to KRAS4b and plays an important role in targeting it to cellular membranes. Note that the farnesyl attachments only penetrate part of the upper leaflet.

Glycosyl-phosphatidylinositol linkers
Normally soluble cytosolic proteins can become attached to membranes through the addition of a glycosyl phophatidylinositol (GPI). The attachment usually contains a conserved tetrasaccharide core of three mannoses (Man) and one unacetylated glucosamine (GlcN) linked to the carboxy terminus of the protein. The GPI can be further modified with extra galactoses and mannoses, as well as additions to the PI group, which secures the protein in the membrane. Figure \(\PageIndex{8}\) shows the common backbone for GPI anchors. Note the additions of the phosphoethanolamines to the core polysaccharide.

GPIs are found in eukaryotic cells and link many surface antigens, adhesion molecules, and hydrolases to the membrane. GPIs from Plasmodium falciparum, the malarial parasite which kills about two million people each year, appear to act as a toxin and are the most common CHO modification of the parasite protein. Mice immunized against the GPI sequence, NH2-CH2-CH2-PO4-Man (α1-2) 6Man (α1-2) Man (α1-6) Man (α1-4) GlcNH2 (α1-6) myo-inositol-1,2-cyclic-phosphate, were substantially protected from malarial symptoms and death after they were exposed to the actual parasite.
Figure \(\PageIndex{9}\) shows a cross-section of a membrane (with cholesterol, PE, SM) containing the glycosylated form of the human complement regulatory protein CD59 protein (1cdr) with a GPI anchor attached at its C-terminus. Note that the middle part of the anchor (glycan) holds the actual protein well above the top of the lipid bilayer. The soluble protein is also glycosylated. The protein binds to complement proteins C8 and/or C9, which are effector immune proteins that assemble on the surface of a cell undergoing lysis.

The GPI anchor is shown in spacefill. Note that it only extends halfway into the bilayer, as you would expect from the size of the fatty acids attached to the phosphatidyl inositol. The glycan part of the GPI is shown in spacefill between the lipid and its protein attachment site. The protein is also glycosylated in the extracellular domain.
A new (5/21) and totally unexpected type of glycosylated molecule has been found at the outer leaflet of mammalian cells - a glycosylated RNA, as shown in Figure \(\PageIndex{10}\). This adds RNA to the lipids and proteins as a target for glycosylation. These surface glycoRNAs interact with antibodies again ds-RNA and the Siglec lectin family. They are found in cells in vivo and cultured cells in vitro.

Figure \(\PageIndex{10}\): A glycoRNA - a small noncoding RNAs with sialylated glycans. Park. https://doi.org/10.14348/molcells.2021.0178
www.molcells.org. Creative Commons Attribution-NonCommercial-ShareAlike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/.
Flynn et al., Small RNAs are modified with N-glycans and displayed on the surface of living cells, Cell (2021), https://doi.org/10.1016/j.cell.2021.04.023
Transmembrane (Integral) Proteins
These proteins pass through the membrane either one in one pass, usually with a single alpha-helix or many membrane-spanning helices. For example, G protein-coupled receptors, often called serpentine receptors, cross the membrane seven times. There are three different types based on the number of types the protein crosses the membrane and the type of secondary structure used in crossing: biotopic (single pass), alpha-helical polytopic, and beta-barrel. These proteins are found in all types of membranes and have many types of functions, from receptors, receptor ligands, structural, adhesion, transport, gene regulation, and transport.
Transmembrane Biotopic - Single Pass Proteins
The are 4 types of single-pass transmembrane proteins:
- Type I: N-terminal outside of the cell (extracellular) and the precursor signal sequence on the N-terminus which is a localization sequence is removed
- Type II: N-terminal intracellular and with the transmembrane domain close to the N-terminus
- Type III: N-terminus extracellular and no signal sequence in precursor protein
- Type IV: N-terminus intracellular and the transmembrane domain close to the C-terminus
The transmembrane domain of single-pass integral membrane proteins consists of a single alpha-helices with nonpolar side chains extending outward from the helical axis where they interact with the nonpolar lipid parts of the membrane. These nonpolar sides are more stable in nonpolar environments.
To study such proteins in a less complex environment, membranes are often "dissolved" in nonpolar, single-chain amphiphilic detergents. These single-chain amphiphiles form micelles in the absence of membrane proteins but can form mixed micelles in which the nonpolar part of the protein is surrounded in the detergent micelle by the nonpolar acyl chains of the detergent.
Figure \(\PageIndex{11}\) (top) below shows just the transmembrane and juxtamembrane (next to the membrane) domains of the single pass Notch protein, which is critical in many signal transduction pathways.

The top images in the figure above show different ways to represent the protein in the bilayer, with the right-hand image showing a cross-section through the membrane to better show how the protein passes through the bilayer.
The bottom images in the figure above show the protein after excess detergent, in this case, octylglucoside, is added to the protein-containing bilayer.
Here are some examples of bitopic single-pass transmembrane proteins
Cadherins
All structures need support and connections. At the macro level, the skeleton supports the mass and organization of organs and tissues in whole organisms. Within an organ, how can cells hold together? How do they adhere to each other? Certainly not through outer leaflet lipid contacts as the outer surface of the leaflet is typically charged. The extracellular matrix does provide some of the glue that holds cells together. At a more detailed level, transmembrane proteins are involved. One class of adhesive proteins is cadherins, a calcium-dependent cell adhesion molecule. There are over 100 human cadherins. They are mostly ditopic, single transmembrane pass proteins. Their cytoplasmic domains interact with proteins like catenin, which then bind to the interior cytoskeletal network composed of actin and other proteins. This provides a way for the intracellular region to regulate the extracellular interactions of the cell.
The extracellular domain is composed of five repeating "cadherin" domains, each around 110 amino acids, that can fold independently. Calcium ions bind at the domain interfaces. A cadherin can interact with other cadherin domains on other cadherins on other cells, leading to cell adhesion. Essentially, the receptor cadherin on one cell binds the ligand cadherin on the other. As metastatic tumor cells lose their adhering feature and leave the site of the primary tumor, you would expect that mutations in cadherins are often involved. They may also be involved in cell sorting during morphogenesis, "regulation of tight and gap junctions, and in the control of intercellular spacing".
Figure \(\PageIndex{12}\) shows a "constructed" image of cadherin-1 (1L3W) interacting with cytoplasmic β-catenin (1I7X) through a modeled transmembrane helix (amino acids QIPAILGILGGILALLILILLLLLFLRR, amino acids 706-731). No full-length structure of cadherin in a membrane is available.

Membrane Protein Kinases
Kinases are enzymes that phosphorylate substrates. Hexokinase is a protein enzyme that catalyzes the phosphorylation of a hexose substrate such as glucose. A protein kinase is a protein enzyme that phosphorylates a protein substrate. That protein could be another copy of itself or another protein. We will see in Chapter 12 that many protein kinases are involved in cell signaling. Many tyrosine protein kinases are bitopic single-pass integral membrane proteins that become active on binding a ligand. Typically, on binding an extracellular ligand, two monomeric copies of the kinase form a dimer in the membrane, activating a tyrosine kinase cytoplasmic domain, which typically phosphorylates (using ATP as a substrate) the other member of the dimer in an "autophosphorylation" reaction. Sometimes the dimers are held together by disulfide bonds.
Figure \(\PageIndex{13}\) shows a "constructed" image of the human dimeric insulin receptor.

One of the monomers is shown in gray. The other monomer is shown in colors corresponding to the domain organization of the protein. Each extracellular dimer (6PXV) has two insulins bound (yellow spacefill). The intracellular domains (1IR3) are activated on insulin binding. No full-length structure of full insulin receptor in a membrane is available.
Almost half of all helical membrane proteins in humans are bitopic, compared to between 20-25% in prokaryotes. Humans have 10-20 fold more bitopic proteins than E.Coli. There appear to be about 196 bitopic proteins in E. coli (located in the inner membrane ) and 70 in M. jannaschii (Archea in plasma membrane). In humans, 57% are in the plasma membrane, with the rest distributed between the Golgi, ER, nuclear, mitochondrial and chloroplast membranes. In single-celled yeast, only 8% are in the plasma membrane.
Beta-Dystroglycan
This protein is another example of a bitopic protein with a single alpha-helix membrane domain. Dystroglycan is a dimer of alpha and beta subunits. Alpha-dystroglycan is a peripheral protein that binds beta-dystroglycan, a transmembrane protein. Alpha dystroglycan also binds lassa virus and lymphocytic choriomeningitis virus glycoprotein, as it serves as viral receptors. It also binds the protein dystrophin, a protein missing in Duchenne muscular dystrophy, which affects 1 out of 5000 live male births. As an integral transmembrane protein, beta-dystroglycan connect the extracellular matrix to the cytoskeleton through dystrophin. Alpha- and beta-dystroglycan share the same gene, which codes one long protein which is proteolyzed post-translationally to form the alpha (N-terminal end) and beta subunits (C-terminal end).
Figure \(\PageIndex{14}\) shows a schematic outline of dystrophin and the dystrophin-associated glycoprotein complex (DAGC).

Figure \(\PageIndex{14}\): Schematic outline of dystrophin and the dystrophin-associated glycoprotein complex (DAGC). Dystrophin contains N-terminal (NT), middle rod,
cysteine-rich (CR), and C-terminal (CT) domains. The middle rod domain is composed of 24 spectrin-like repeats (numerical numbers in the cartoon, positively
charged repeats are marked in white color) and four hinges (H1, H2, H3, and H4). Dystrophin has two actin-binding domains located at NT and repeats 11-15,
respectively. Repeats 1-3 interact with the negatively charged lipid bilayer. Repeats 16 and 17 form the neuronal nitric oxide synthase (nNOS)-binding domain.
Dystrophin interacts with microtubules through repeats 20-23. Part of H4 and the CR domain binds to the β-subunit of dystroglycan (βDG). The CT domain of
dystrophin interacts with syntrophin (Syn) and dystrobrevin (Dbr). Dystrophin links components of the cytoskeleton (actin and microtubule) to laminin in the
extracellular matrix. Sarcoglycans and sarcospan do not interact with dystrophin directly but they strengthen the entire DAGC, which consists of dystrophin,
DG, sarcoglycans, sarcospan, Syn, Dbr, and nNOS. .Disease Models & Mechanisms (2015) doi:10.1242/dmm.018424vailable via license: CC BY 3.0
Transmembrane - Alpha-helical polytopic
There are so many intriguing examples of these proteins. We'll illustrate just two.
Rhodopsin-like receptors and pumps
These proteins are involved in cell signaling and are the target of most pharmaceutic drugs. G protein-coupled receptors (GPCRs) are incredibly important and we will discuss them extensively in Chapter 12.
GPCRs are cell receptors that span the membrane seven times in a serpentine fashion. They bind ligands (neurotransmitters, hormones, etc) in the extracellular or internal membrane domains (the latter for hydrophobic ligands), and through propagated conformations changes alter the cytoplasmic domain where they functionally interact with a heterotrimeric G protein.
Figure \(\PageIndex{15}\) shows an interactive iCn3D model of the human cannabinoid receptor with bound cholesterol and Δ 9 -tetrahydrocannabinol (Δ9 -THC) in spacefill (5xra). The red dummy atoms represent the outer leaflet and the blue the inner.
.png?revision=1&size=bestfit&width=275&height=288)
Δ 9 -THC is a partial agonist and tunes the response of the receptor. The active site is conformationally somewhat flexible or plastic. Other ligands bound to it act as antagonists instead of agonists and must do so by eliciting nonactive conformations.
ABC Transporter
Figure \(\PageIndex{16}\) shows an interactive iCn3D model of the P-glycoprotein multidrug resistance transporter protein (6nf1).
.png?revision=1)
The spacefill ligands represent Zosuquidar, which binds with high affinity to P-glycoprotein and inhibits its activity, making it a cancer agent as it prevents chemotherapeutic drugs that have entered the cell from being pumped out. The protein chain interacting with it on the cytoplasmic face is an antibody fragment used to stabilize the P-glycoprotein so crystals could form.
Transmembrane Beta-barrel transmembrane
We will focus on two of these proteins.
Outer Membrane Factor (OMF) - Gram-negative bacteria
Figure \(\PageIndex{17}\) shows an interactive iCn3D model of a beta-barrel transmembrane protein OPRM - Outer Membrane Factor (4y1k) from Pseudomonas aeruginosa that acts as a pore. It also has a palmitoyl fatty acid in thioester linkage to Cys 1 of the protein for extra but unneeded anchorage.
%25C2%25A0.png?revision=1&size=bestfit&width=245&height=336)
Use this link for another view: https://structure.ncbi.nlm.nih.gov/i...FtY5FL7&t=4Y1K (OPM) in iCn3D
This protein is part of a large complex of proteins that spans both the inner and outer membranes of Gram-negative (examples E. Coli and Pseudomonas aeruginosa) bacterial cell walls. Unfortunately for humans, this protein complex pumps out toxins (to the bacteria) like antibiotics, which makes bacteria resistant to these drugs. The OPRM acts as the outer passageway or duct for the pumped molecules. The Bacterial Outer Membrane Factor (OMF) protein differs in sequence but all form the beta-barrel duct. The E. Coli version of OMF has a triacylated lipid modification of the N-terminus. The N terminal lipid modification might be necessary for the initial attachment of the protein to a membrane before the insertion of the beta-barrel. As such, the enzymes involved in the attachment of the tail could be targets for new antibiotics.
Voltage-dependent anion channel (VDAC) - mouse
This protein regulates the movement of molecules between the cytoplasm and the interior of the mitochondria across the outer mitochondrial membrane. VDAC also serves as a docking site or scaffold for the assembly of molecules into a complex that regulates mitochondrial function. The protein's conformation and hence function are regulated by changes in the transmembrane potential, which we will explore in the next sections. Hence the protein and its function are voltage-dependent. Figure \(\PageIndex{18}\) shows an interactive iCn3D model of mouse VDAC with a beta-barrel formed by 19 beta-strands (3emn).
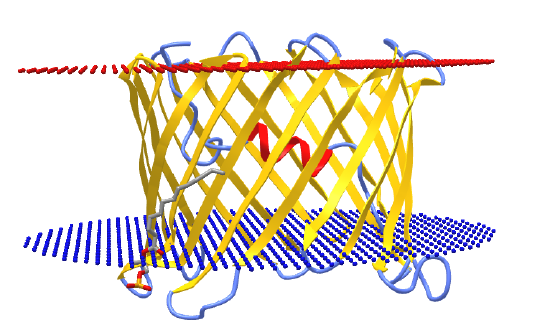
Note the N-terminal alpha-helix resident inside the channel opening. This helical section moves on changes in membrane potential, gating open and hence regulating the flow of metabolites and ions across the membrane through the pore. At a low transmembrane potential (10 mV), the conductance is high as the channel is in the open state. When the potential increases to 30 mV (either + or -) conductance drops as the protein forms the closed state.
Now that you understand the structure of membrane proteins, let's explore a key type of function of a subset of integral membrane proteins: the movement of molecules/ions across the membrane.