
1.1: Cellular Foundations
- Page ID
- 102240


\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)(Learning goals written by Claude, Sonnet 4.6, Anthropic)
Cells as Chemical Systems
- Describe how a cell functions analogously to a chemical factory, distinguishing between anabolic and catabolic processes and explaining how metabolic pathways integrate both.
- Explain how enzymes accelerate metabolic reactions and regulate pathway flux, and contrast the lock-and-key and induced-fit models of substrate binding.
Molecular Traffic Across Membranes
- Compare the mechanisms by which small molecules, ions, and large particles cross cellular membranes, distinguishing passive diffusion, facilitated transport, active transport, ion channels, pores, and endocytosis/exocytosis.
- Explain how the amphiphilic nature of the lipid bilayer determines which molecules require protein-mediated transport.
Cellular Organization and the Intracellular Environment
- Identify the major organelles of a eukaryotic animal cell and assign each a primary biochemical function, noting how compartmentalization separates anabolic from catabolic processes.
- Describe the composition and roles of the three cytoskeletal elements and explain how motor proteins use cytoskeletal tracks to direct intracellular cargo transport.
- Explain how molecular crowding in the cytoplasm affects enzyme activity, protein stability, and the localization of metabolic functions, and discuss why in vitro biochemical studies may not fully replicate in vivo conditions.
- Explain how phase transitions and liquid-liquid demixing drive the formation of subcellular structures such as lipid droplets, membrane rafts, and RNA-protein particles, and identify the noncovalent interactions underlying these phenomena.
Introduction
You have probably studied the cell many times, either in high school or in college biology classes. Many websites review prokaryotic (bacterial and archaeal) and eukaryotic cells (including protists, fungi, plants, and animals). All cells share similar structural components, including genetic material in the form of chromosomes, a membrane-bound lipid bilayer that separates the cell's interior from its exterior, and ribosomes responsible for protein synthesis. This tutorial is designed specifically from a chemistry perspective. It explores four classes of biomolecules that are also present in all cell types (lipids, proteins, nucleic acids, and carbohydrates) and describes, in a simplified pictorial manner, where they are found, produced, and degraded in a typical eukaryotic animal cell (i.e., their lifecycle). This cell review focuses on the organelle structures common in eukaryotic cells. Subsequent chapters will concentrate on the structure and function of specific biomolecules.
Let’s think of a cell as a chemical factory that designs, imports, synthesizes, uses, exports, and degrades various chemicals (in the case of the cell, these include lipids, proteins, nucleic acids, and carbohydrates). It must also determine or sense the amount of raw and finished chemicals available and respond to its own and external needs by adjusting production accordingly. Biochemistry is the branch of science that studies the chemical processes within a cell. Understanding these processes can also lend insight into disease states and the pharmacological effects of toxins, drugs, and other medicines within the body.
The building and breaking down of life-sustaining chemicals within an organism is known as Metabolism. Overall, metabolism involves: (1) the net exergonic, energy-releasing metabolism of food/fuel to power endergonic, energy-requiring cellular processes; (2) the conversion of food/fuel to building blocks for the synthesis of proteins, lipids, nucleic acids, and other biomolecules; and (3) the elimination of waste products. These processes enable organisms to grow, reproduce, maintain their structures, and adapt to their environments.
Metabolic reactions may be categorized as catabolic– the breaking down of compounds (for example, the breaking down of proteins into amino acids during digestion)- or anabolic – the building up (synthesis) of compounds (such as proteins, carbohydrates, lipids, and nucleic acids). Typically, catabolism releases energy, while anabolism consumes it.
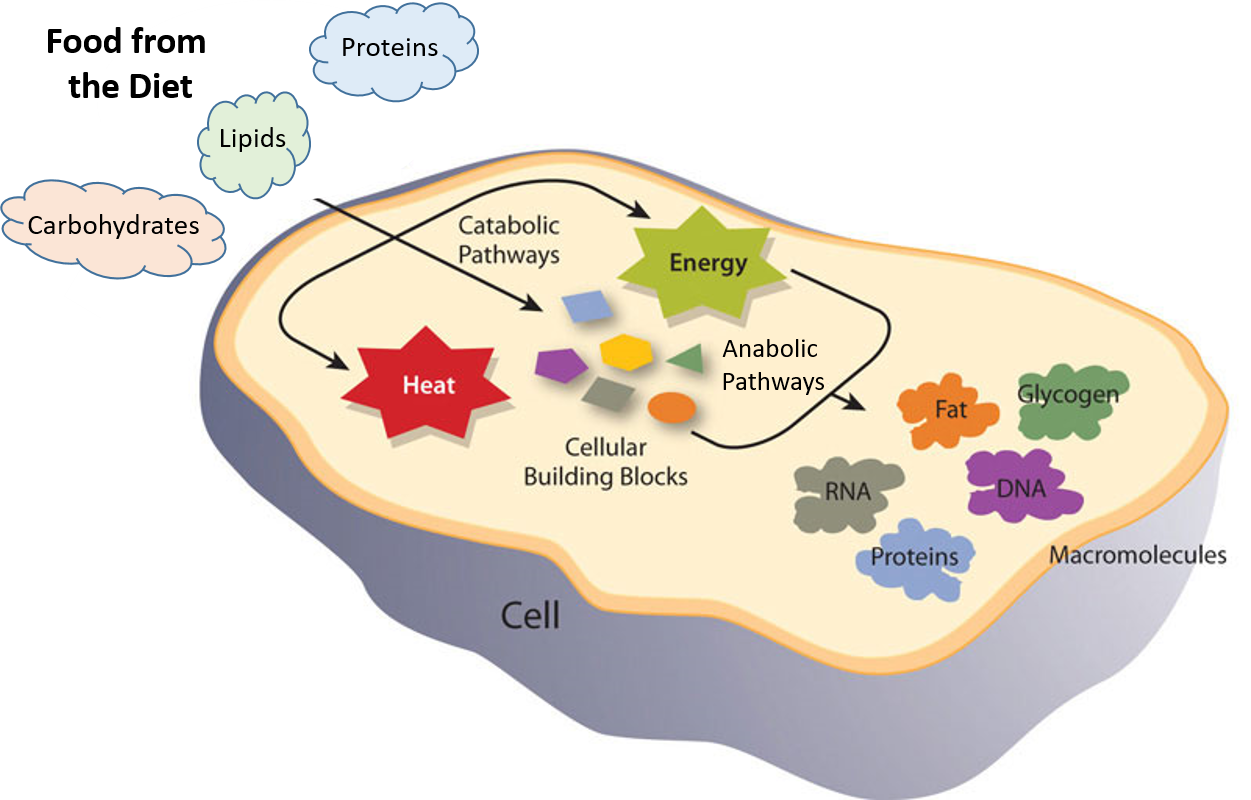
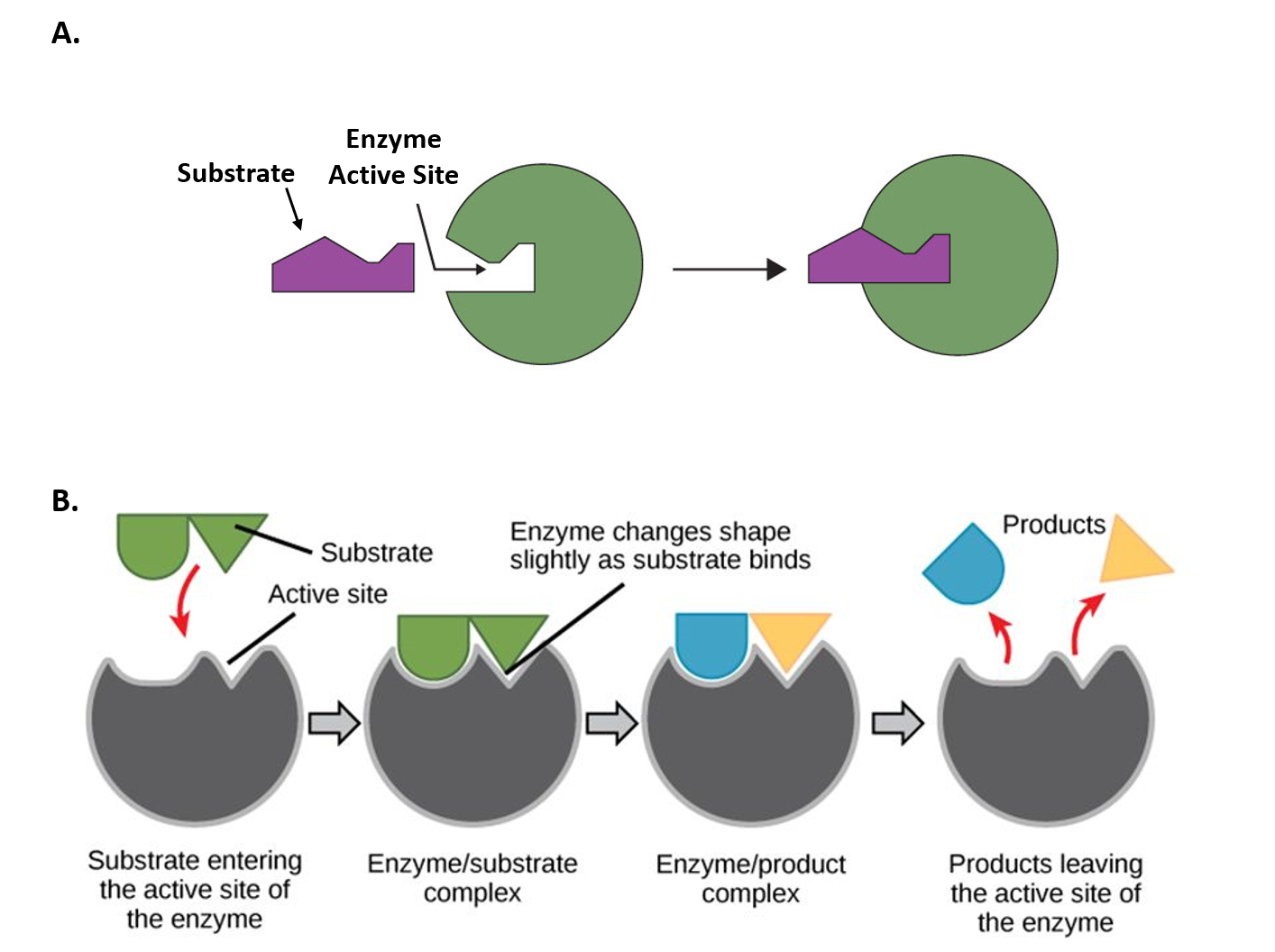
The chemical reactions of metabolism are organized into metabolic pathways, in which one chemical is transformed through a series of steps into another, each step often catalyzed by a specific enzyme. Enzymes are crucial to metabolism because they act as catalysts, allowing reactions to proceed more rapidly. Additionally, enzymes can provide a mechanism for cells to regulate the rate of a metabolic reaction in response to changes in the cell’s environment or to signals from other cells by activating or inhibiting the enzyme’s activity. Enzymes can also enable organisms to drive desirable reactions that require energy by coupling them to spontaneous reactions that release energy. Enzyme shape is critical to enzyme function, as it determines the specific binding of a reactant. This can occur through a lock-and-key model, in which the reactant fits the exact shape of the enzyme's binding site, or through an induced-fit model, in which contact between the reactant and the protein changes the protein's shape, allowing it to bind. The catalytic mechanisms, kinetics, and regulatory pathways of enzymes will be studied in detail within this text.
Within eukaryotic cells, the metabolic machinery enables the construction of membrane-bound organelles that help compartmentalize cellular functions. Therefore, organelles, having discrete cellular functions, can be considered ‘little organs’ within the cell. The figure of the cell below and on the other linked sites was made available with Liliana Torres's kind permission. For more detailed information, click on the blue hyperlinks for some organelles.
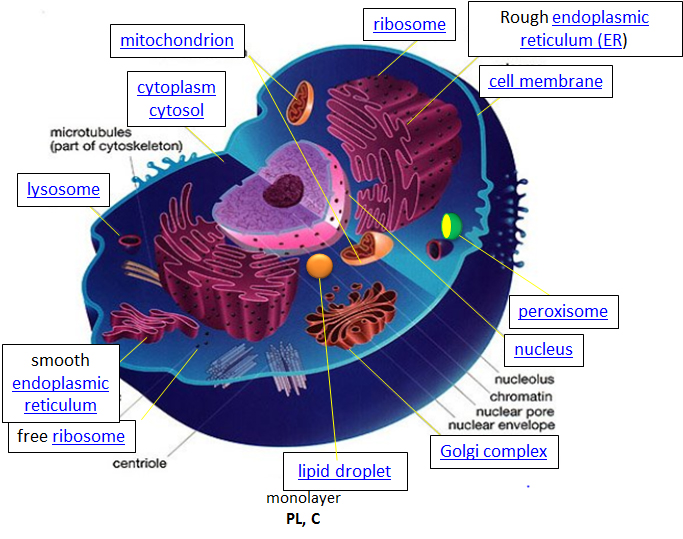
Design – The design for a cell primarily resides in the blueprint for the cell, the genetic code, which comprises the DNA in the cell nucleus and a small amount in the mitochondria. The DNA blueprint must be read out (transcribed) by DNA readers (DNA-dependent RNA polymerases) to form RNA. Since DNA and RNA are nucleic acids composed of deoxynucleotides (in DNA) and nucleotides (in RNA), this process is called transcription. A type of RNA, messenger RNA, is then decoded to form a new polymer, a protein composed of amino acid monomers. This process is called translation (analogous to converting an English sentence into a Spanish one). In a nanomachine, the ribosome, which contains RNA and protein subunits, interacts with the messenger RNA and incoming transfer RNA, which is connected to individual amino acids, to create a protein. The genetic code has a master plan that determines the sequence of all cellular proteins, catalyzing nearly all cellular activities, including catalysis, motility, and the formation of architectural structures. In contrast to DNA, RNA, and protein polymers, the length and sequence of polysaccharide polymers and lipids are not determined by a template, but rather by the enzymes that catalyze their synthesis.
Import/Export: Many of the chemical constituents of the cell arise not from direct synthesis but from the import of both small and large molecules. The imported molecules must pass through the cell membrane and, in some cases, through additional membranes if they need to reside inside membrane-bound organelles. Molecules can move into the cell by passive diffusion across the membrane, but their movement is usually “facilitated” by a membrane transporter protein. Molecules can also move against a concentration gradient in a process called “active transport.” Given the amphiphilic nature of the bilayer (polar head group exterior, nonpolar interior), you would expect that polar molecules like glucose would have difficulty moving across the membrane by passive diffusion. Typically, only small nonpolar molecules move across the membrane via passive transport. Membrane-bound transport proteins mediate the movement of both nonpolar and polar molecules.
- transporters, carrier proteins, and permeases: These membrane proteins facilitate the movement of specific ligand molecules across a membrane, typically down a concentration gradient. Click the image below for a computer (molecular dynamics) simulation of the facilitated diffusion of lactose across the membrane by a membrane protein (lactose permease).

Morten Ø. Jensen et al. Sugar Transport across Lactose Permease Probed by Steered Molecular Dynamics. Open Archive. DOI:https://doi.org/10.1529/biophysj.107.103994. Theoretical and Computational Biophysics Group, Beckman Institute, Urbana, Illinois, USA. Department of Physics, University of Illinois at Urbana-Champaign, Urbana, Illinois USA. Creative Commons licensing
https://www.cell.com/cms/10.1529/bio...329ed/mmc2.mp4
- ion channels: These membrane proteins allow the flow of ions across membranes. Some are permanently open (nongated), while others are gated, opening or closing depending on the presence of ligands that bind to the protein channel and the local environment of the protein in the membrane. The flow of ions through the channel proceeds in a thermodynamically favored direction, determined by the concentration and voltage gradients across the membrane.

Fatemeh Khalili-Araghi et al. Dynamics of K+ Ion Conduction through Kv1.2. Open Archive DOI:https://doi.org/10.1529/biophysj.106.091926. Theoretical and Computational Biophysics Group, Beckman Institute, Urbana, Illinois, USA. Department of Physics, University of Illinois at Urbana-Champaign, Urbana, Illinois
- Pores: Some membranes (nuclear, mitochondrial) assemble proteins (such as porins) to form large but regulated pores. Porins are found in mitochondrial membranes, while nucleoporins are found in the nuclear membrane. Small molecules can generally pass through these membrane pores, while large ones are selected based on their tendency to form transient noncovalent attractive forces with the proteins in the pore. The following link shows the diffusion of water through aquaporin. Animation of water diffusion through the aquaporin channel,

Water Transport in Aquaporins. https://www.ks.uiuc.edu/Research/aquaporins/. Theoretical and Computational Biophysics Group, Beckman Institute, Urbana, Illinois, USA. Department of Physics, University of Illinois at Urbana-Champaign, Urbana, Illinois
- Endocytosis: Very large particles [for example, Low-Density Lipoproteins (LDL) and viruses] can enter a cell through endocytosis. Initially, the LDL or virus binds to a receptor on the cell's surface. This triggers a series of events that lead to the invagination of the cell membrane at that point. This eventually pinches off to form an endosomal vesicle surrounded by clathrin, a protein. “Early” endosomes can pick up new proteins and other constituents and shed them as they move and mature through the cell. During this maturation process, protein pumps in the endosome decrease endosomal pH, which can result in conformational changes in protein structure and protein shedding. Eventually, the “late” endosome reaches and fuses with the lysosome, an internal organelle that contains enzymes for degradation. Undegraded components, such as viral nucleic acids or cholesterol, are delivered to the cell. This transport can also go in the reverse direction (called exocytosis) and recycle receptors to the cell membrane. Likewise, vesicles pinched off from the Golgi complex can fuse with endosomes, with some components surviving the process to reenter the Golgi.

Alexey Solodovnikov (Idea, Producer, CG, Editor), Valeria Arkhipova (Scientific Сonsultant). Endocytosis. For example, coronavirus binds to the ACE2 receptor of the epithelial cell. CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
Synthesize/Degrade: Cells must synthesize and degrade small molecules, as well as larger polymers such as proteins, carbohydrates, lipids, and nucleic acids. The anabolic (synthetic) and catabolic (degradative) pathways are often compartmentalized in time and space within a cell. For example, fatty acid synthesis occurs in the cytoplasm, whereas fatty acid oxidation takes place in the mitochondria. Proteins are synthesized in the cytoplasm or completed in the endoplasmic reticulum (for membrane and exported proteins). They are degraded in the lysosome or, more importantly, in a large multimolecular structure in the cell called the proteasome.
Key Characteristics of a Cell
Let’s consider some key cell characteristics as a prelude to later chapters.
Cells and their internal compartments have regulated concentrations of ions and hydronium ions.
As expected, the pH of the cytosol (the aqueous substance surrounding all the organelles within the cell) varies from about 7.0 to 7.4, depending on the metabolic state of the cell. Some organelles contain proton transporters that can significantly alter their pH. For example, the pH inside the lysosome, a degradative organelle, is about 4.8. Furthermore, creating a pH gradient across the inner mitochondrial membrane is sufficient to drive the thermodynamically unfavored synthesis of ATP.
Compared to the extracellular fluid, the concentration of potassium ions is higher inside the cell. In contrast, sodium, chloride, and calcium ion concentrations are higher outside the cell (see table below). Ion transporters and channels maintain these concentration gradients, requiring energy expenditure — ultimately ATP hydrolysis. Changes in these concentrations are integral to the signaling system that the cell uses to sense and respond to changes in its external and internal environments. The table below shows approximate ion concentrations in the cell.
| Ion | Inside (mM) | Outside (mM) |
|---|---|---|
| Na+ | 140 | 5 |
| K+ | 12 | 140 |
| Cl- | 4 | 15 |
| Ca2+ | 1 uM | 2 |
Cells have an internal framework that provides architectural and internal structural support.
The “cytoskeletal” architecture of a cell (with molecular “cables” and “girder-like” structures) is superficially similar to that of a factory. The cell's internal framework, the cytoskeleton, is composed of microfilaments, intermediate filaments, and microtubules. These, in turn, are built from proteins that self-assemble to form the internal architecture. Parts of the cytoskeleton can be seen in Figure 1.4.

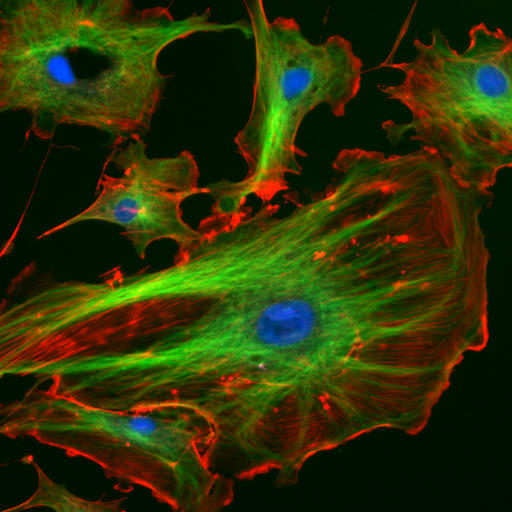
Microfilaments of actin monomers (stained with a red/orange fluorophore) and microtubules, which offer more structural support made of tubulin monomers (stained green), and the blue-stained nucleus are shown in the image. Organelles are supported and organized by the cytoskeleton (primarily microtubules). Actin (stained orange) and spectrin microfilaments support the cell membrane underneath the inner leaflet. Motor proteins like myosin (that moves along actin microfilaments) and dynein and kinesin (that move along tubulin microtubules) carry cargo (vesicles, organelles) directionally. The cell is not a disorganized collection of molecules and organelles. Instead, it is highly organized to optimize chemical production, use, and degradation.
Cells have various shapes. Some circulating immune cells must slip between the cells lining capillary walls to migrate to sites of infection. The same process occurs when tumor cells metastasize and spread to other parts of the body. To do so, the cell must drastically change shape, a response that requires dissociation of cytoskeletal polymers into monomers, which are later available for polymerization. The following video shows the mobility and flexibility of a Killer T-cell as it attacks and kills a cancerous cell.
Video 1.1 Killer T Cell Attacking Cancer. Video available on YouTube through Creative Commons by Cambridge University
The cell is an amazingly crowded place.
In chemistry labs, we typically work with dilute solutions of solute molecules in a solvent. You have probably heard that the body is composed of approximately 68% water, but the water concentration varies depending on the cellular environment. Solute molecules, such as proteins and carbohydrates, are densely packed. Cells are so crowded that the space between larger molecules like proteins is typically smaller than that of a single protein. Studies have shown that protein stability increases under these conditions, helping keep it in its correctly folded, native state. Another consequence of high intracellular concentrations is that it limits the diffusion of molecules throughout the cell, as expected from an equilibrium perspective in dilute solutions. Thus, cytoplasmic cellular functions can be highly localized within specific cell regions, creating unique microenvironments and higher differentiation potential within a single cell.
Hence, the study of biomolecules in dilute solutions in the lab may not capture the full complexity of their interactions and activities in vivo. Recently, investigators have added a neutral copolymer of sucrose and epichlorohydrin to cells in vitro. These particles organized extracellular molecules secreted by the cell, forming an extracellular “matrix” that organized the microfilaments on the inside and altered cell activity.1 Furthermore, in vitro enzyme activity of a key enzyme in glycolysis dramatically increases under crowded conditions.2 Another result of crowding may be the spatial and temporal association of key enzymes involved in specific metabolic pathways, allowing for the coordinated passage of substrates and products within the colocalized enzyme system.
Cell components undergo phase transitions to form substructures within the cell.
A perplexing question is how substructures form within a cell. This includes the biogenesis of organelles, such as mitochondria, and smaller particles, including polysaccharide granules, lipid droplets, protein/RNA particles (including ribosomes), and the nucleolus of the cell nucleus. It might be easiest to consider this problem using two examples from the lipid world: lipid droplets and membrane rafts. Phase transitions occur when a sparingly soluble nonpolar liquid is added to water. At a high enough concentration, the solubility of the nonpolar liquid is exceeded, and a phase transition occurs, as evidenced by the appearance of two separate liquid phases. The same process occurs when triglycerides coalesce into lipid droplets with proteins associated with their surface. Another example occurs within a cell membrane, where lipids with saturated alkyl chains self-associate with membrane cholesterol (which contains a rigid, planar ring system) to form a lipid raft microdomain. Lipid rafts are characterized by greater packing efficiency, rigidity, and thickness than other membrane parts. These lipid rafts often recruit proteins involved in signaling processes within the cell membranes. This phase separation process is known as liquid-liquid demixing, in which two “liquid-like” substances separate.
Similarly, it appears that proteins that interact with RNA are composed of less diverse amino acid sequences and have more flexible (“more liquid-like) structures, allowing their preferential interaction with RNA to form large RNA-protein particles (like the ribosome and other RNA processing structures) in a fashion that mimics liquid/liquid demixing. All of these interactions are just manifestations of the various noncovalent interactions that can exist between molecules. These include ionic, ion-dipole, dipole-dipole, and London dispersion force interactions. A review of noncovalent interactions can be found in a Khan Academy video on YouTube.
Summary
(Summary written by Claude, Sonnet 4.6, Anthropic)
This chapter introduces the cell as an integrated chemical system and establishes the conceptual foundation for the biochemistry that follows. Rather than reviewing cell biology descriptively, it frames cellular organization through the lens of chemistry and chemical logic.
Metabolism — the sum of all chemical transformations within a cell — is organized into catabolic pathways that release energy by breaking down molecules, and anabolic pathways that consume energy to synthesize them. These reactions are catalyzed by enzymes, which accelerate reactions, couple energetically unfavorable processes to favorable ones, and provide the cell with exquisite regulatory control. Enzyme–substrate recognition occurs either through rigid complementarity (lock-and-key) or through conformational adaptation upon binding (induced fit).
The eukaryotic cell achieves biochemical efficiency through compartmentalization: distinct organelles carry out discrete functions, separating, for example, fatty acid synthesis (cytoplasm) from fatty acid oxidation (mitochondria), and protein synthesis (ribosomes/ER) from protein degradation (lysosomes/proteasome). The genetic blueprint residing in nuclear DNA directs protein synthesis through the sequential processes of transcription and translation, with the ribosome serving as the central nanomachine for decoding messenger RNA.
Molecular traffic across membranes is tightly controlled. Because the lipid bilayer's nonpolar interior restricts the passage of polar and charged molecules, cells rely on transporters, ion channels, aquaporins, and nuclear pores to regulate molecular movement. Large particles enter and exit via endocytosis and exocytosis, processes that also recycle receptors and redistribute membrane components.
Three additional physical properties of the cell have important biochemical consequences. First, ion concentration gradients — maintained at energetic cost — power signaling events and ATP synthesis. Second, the cytoskeleton (actin microfilaments, intermediate filaments, and tubulin microtubules) provides structural support and serves as a directional highway system for motor proteins transporting organelles and vesicles. Third, the molecular crowding of the cytoplasm — far denser than typical laboratory solutions — enhances protein stability, restricts free diffusion, and promotes the spatial clustering of enzymes within shared pathways. Finally, phase separation through liquid-liquid demixing underlies the spontaneous formation of lipid droplets, membrane microdomains (rafts), and RNA-protein assemblies, all driven by the noncovalent interactions that will recur throughout this text.