
23.1: Eukaryotic Origins
- Page ID
- 1939

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Skills to Develop
- List the unifying characteristics of eukaryotes
- Describe what scientists know about the origins of eukaryotes based on the last common ancestor
- Explain endosymbiotic theory
Living things fall into three large groups: Archaea, Bacteria, and Eukarya. The first two have prokaryotic cells, and the third contains all eukaryotes. A relatively sparse fossil record is available to help discern what the first members of each of these lineages looked like, so it is possible that all the events that led to the last common ancestor of extant eukaryotes will remain unknown. However, comparative biology of extant organisms and the limited fossil record provide some insight into the history of Eukarya.
The earliest fossils found appear to be Bacteria, most likely cyanobacteria. They are about 3.5 billion years old and are recognizable because of their relatively complex structure and, for prokaryotes, relatively large cells. Most other prokaryotes have small cells, 1 or 2 µm in size, and would be difficult to pick out as fossils. Most living eukaryotes have cells measuring 10 µm or greater. Structures this size, which might be fossils, appear in the geological record about 2.1 billion years ago.
Characteristics of Eukaryotes
Data from these fossils have led comparative biologists to the conclusion that living eukaryotes are all descendants of a single common ancestor. Mapping the characteristics found in all major groups of eukaryotes reveals that the following characteristics must have been present in the last common ancestor, because these characteristics are present in at least some of the members of each major lineage.
- Cells with nuclei surrounded by a nuclear envelope with nuclear pores. This is the single characteristic that is both necessary and sufficient to define an organism as a eukaryote. All extant eukaryotes have cells with nuclei.
- Mitochondria. Some extant eukaryotes have very reduced remnants of mitochondria in their cells, whereas other members of their lineages have “typical” mitochondria.
- A cytoskeleton containing the structural and motility components called actin microfilaments and microtubules. All extant eukaryotes have these cytoskeletal elements.
- Flagella and cilia, organelles associated with cell motility. Some extant eukaryotes lack flagella and/or cilia, but they are descended from ancestors that possessed them.
- Chromosomes, each consisting of a linear DNA molecule coiled around basic (alkaline) proteins called histones. The few eukaryotes with chromosomes lacking histones clearly evolved from ancestors that had them.
- Mitosis, a process of nuclear division wherein replicated chromosomes are divided and separated using elements of the cytoskeleton. Mitosis is universally present in eukaryotes.
- Sex, a process of genetic recombination unique to eukaryotes in which diploid nuclei at one stage of the life cycle undergo meiosis to yield haploid nuclei and subsequent karyogamy, a stage where two haploid nuclei fuse together to create a diploid zygote nucleus.
- Members of all major lineages have cell walls, and it might be reasonable to conclude that the last common ancestor could make cell walls during some stage of its life cycle. However, not enough is known about eukaryotes’ cell walls and their development to know how much homology exists among them. If the last common ancestor could make cell walls, it is clear that this ability must have been lost in many groups.
Endosymbiosis and the Evolution of Eukaryotes
In order to understand eukaryotic organisms fully, it is necessary to understand that all extant eukaryotes are descendants of a chimeric organism that was a composite of a host cell and the cell(s) of an alpha-proteobacterium that “took up residence” inside it. This major theme in the origin of eukaryotes is known as endosymbiosis, one cell engulfing another such that the engulfed cell survives and both cells benefit. Over many generations, a symbiotic relationship can result in two organisms that depend on each other so completely that neither could survive on its own. Endosymbiotic events likely contributed to the origin of the last common ancestor of today’s eukaryotes and to later diversification in certain lineages of eukaryotes (Figure \(\PageIndex{4}\)). Before explaining this further, it is necessary to consider metabolism in prokaryotes.
Prokaryotic Metabolism
Many important metabolic processes arose in prokaryotes, and some of these, such as nitrogen fixation, are never found in eukaryotes. The process of aerobic respiration is found in all major lineages of eukaryotes, and it is localized in the mitochondria. Aerobic respiration is also found in many lineages of prokaryotes, but it is not present in all of them, and many forms of evidence suggest that such anaerobic prokaryotes never carried out aerobic respiration nor did their ancestors.
While today’s atmosphere is about one-fifth molecular oxygen (O2), geological evidence shows that it originally lacked O2. Without oxygen, aerobic respiration would not be expected, and living things would have relied on fermentation instead. At some point before, about 3.5 billion years ago, some prokaryotes began using energy from sunlight to power anabolic processes that reduce carbon dioxide to form organic compounds. That is, they evolved the ability to photosynthesize. Hydrogen, derived from various sources, was captured using light-powered reactions to reduce fixed carbon dioxide in the Calvin cycle. The group of Gram-negative bacteria that gave rise to cyanobacteria used water as the hydrogen source and released O2 as a waste product.
Eventually, the amount of photosynthetic oxygen built up in some environments to levels that posed a risk to living organisms, since it can damage many organic compounds. Various metabolic processes evolved that protected organisms from oxygen, one of which, aerobic respiration, also generated high levels of ATP. It became widely present among prokaryotes, including in a group we now call alpha-proteobacteria. Organisms that did not acquire aerobic respiration had to remain in oxygen-free environments. Originally, oxygen-rich environments were likely localized around places where cyanobacteria were active, but by about 2 billion years ago, geological evidence shows that oxygen was building up to higher concentrations in the atmosphere. Oxygen levels similar to today’s levels only arose within the last 700 million years.
Recall that the first fossils that we believe to be eukaryotes date to about 2 billion years old, so they appeared as oxygen levels were increasing. Also, recall that all extant eukaryotes descended from an ancestor with mitochondria. These organelles were first observed by light microscopists in the late 1800s, where they appeared to be somewhat worm-shaped structures that seemed to be moving around in the cell. Some early observers suggested that they might be bacteria living inside host cells, but these hypotheses remained unknown or rejected in most scientific communities.
Endosymbiotic Theory
As cell biology developed in the twentieth century, it became clear that mitochondria were the organelles responsible for producing ATP using aerobic respiration. In the 1960s, American biologist Lynn Margulis developed endosymbiotic theory, which states that eukaryotes may have been a product of one cell engulfing another, one living within another, and evolving over time until the separate cells were no longer recognizable as such. In 1967, Margulis introduced new work on the theory and substantiated her findings through microbiological evidence. Although Margulis’ work initially was met with resistance, this once-revolutionary hypothesis is now widely (but not completely) accepted, with work progressing on uncovering the steps involved in this evolutionary process and the key players involved. Much still remains to be discovered about the origins of the cells that now make up the cells in all living eukaryotes.
Broadly, it has become clear that many of our nuclear genes and the molecular machinery responsible for replication and expression appear closely related to those in Archaea. On the other hand, the metabolic organelles and genes responsible for many energy-harvesting processes had their origins in bacteria. Much remains to be clarified about how this relationship occurred; this continues to be an exciting field of discovery in biology. For instance, it is not known whether the endosymbiotic event that led to mitochondria occurred before or after the host cell had a nucleus. Such organisms would be among the extinct precursors of the last common ancestor of eukaryotes.
Mitochondria
One of the major features distinguishing prokaryotes from eukaryotes is the presence of mitochondria. Eukaryotic cells may contain anywhere from one to several thousand mitochondria, depending on the cell’s level of energy consumption. Each mitochondrion measures 1 to 10 or greater micrometers in length and exists in the cell as an organelle that can be ovoid to worm-shaped to intricately branched (Figure \(\PageIndex{1}\)). Mitochondria arise from the division of existing mitochondria; they may fuse together; and they may be moved around inside the cell by interactions with the cytoskeleton. However, mitochondria cannot survive outside the cell. As the atmosphere was oxygenated by photosynthesis, and as successful aerobic prokaryotes evolved, evidence suggests that an ancestral cell with some membrane compartmentalization engulfed a free-living aerobic prokaryote, specifically an alpha-proteobacterium, thereby giving the host cell the ability to use oxygen to release energy stored in nutrients. Alpha-proteobacteria are a large group of bacteria that includes species symbiotic with plants, disease organisms that can infect humans via ticks, and many free-living species that use light for energy. Several lines of evidence support that mitochondria are derived from this endosymbiotic event. Most mitochondria are shaped like alpha-proteobacteria and are surrounded by two membranes, which would result when one membrane-bound organism was engulfed into a vacuole by another membrane-bound organism. The mitochondrial inner membrane is extensive and involves substantial infoldings called cristae that resemble the textured, outer surface of alpha-proteobacteria. The matrix and inner membrane are rich with the enzymes necessary for aerobic respiration.
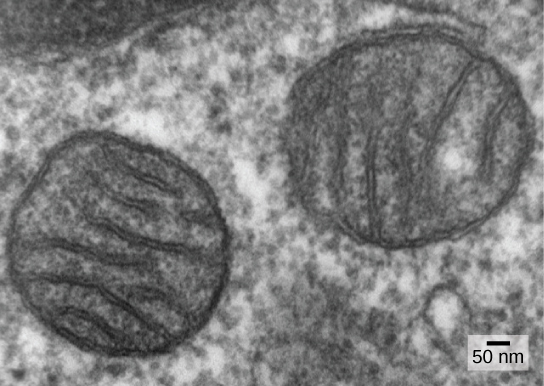
Mitochondria divide independently by a process that resembles binary fission in prokaryotes. Specifically, mitochondria are not formed from scratch (de novo) by the eukaryotic cell; they reproduce within it and are distributed with the cytoplasm when a cell divides or two cells fuse. Therefore, although these organelles are highly integrated into the eukaryotic cell, they still reproduce as if they are independent organisms within the cell. However, their reproduction is synchronized with the activity and division of the cell. Mitochondria have their own (usually) circular DNA chromosome that is stabilized by attachments to the inner membrane and carries genes similar to genes expressed by alpha-proteobacteria. Mitochondria also have special ribosomes and transfer RNAs that resemble these components in prokaryotes. These features all support that mitochondria were once free-living prokaryotes.
Mitochondria that carry out aerobic respiration have their own genomes, with genes similar to those in alpha-proteobacteria. However, many of the genes for respiratory proteins are located in the nucleus. When these genes are compared to those of other organisms, they appear to be of alpha-proteobacterial origin. Additionally, in some eukaryotic groups, such genes are found in the mitochondria, whereas in other groups, they are found in the nucleus. This has been interpreted as evidence that genes have been transferred from the endosymbiont chromosome to the host genome. This loss of genes by the endosymbiont is probably one explanation why mitochondria cannot live without a host.
Some living eukaryotes are anaerobic and cannot survive in the presence of too much oxygen. Some appear to lack organelles that could be recognized as mitochondria. In the 1970s to the early 1990s, many biologists suggested that some of these eukaryotes were descended from ancestors whose lineages had diverged from the lineage of mitochondrion-containing eukaryotes before endosymbiosis occurred. However, later findings suggest that reduced organelles are found in most, if not all, anaerobic eukaryotes, and that all eukaryotes appear to carry some genes in their nuclei that are of mitochondrial origin. In addition to the aerobic generation of ATP, mitochondria have several other metabolic functions. One of these functions is to generate clusters of iron and sulfur that are important cofactors of many enzymes. Such functions are often associated with the reduced mitochondrion-derived organelles of anaerobic eukaryotes. Therefore, most biologists accept that the last common ancestor of eukaryotes had mitochondria.
Plastids
Some groups of eukaryotes are photosynthetic. Their cells contain, in addition to the standard eukaryotic organelles, another kind of organelle called a plastid. When such cells are carrying out photosynthesis, their plastids are rich in the pigment chlorophyll a and a range of other pigments, called accessory pigments, which are involved in harvesting energy from light. Photosynthetic plastids are called chloroplasts (Figure \(\PageIndex{2}\)).
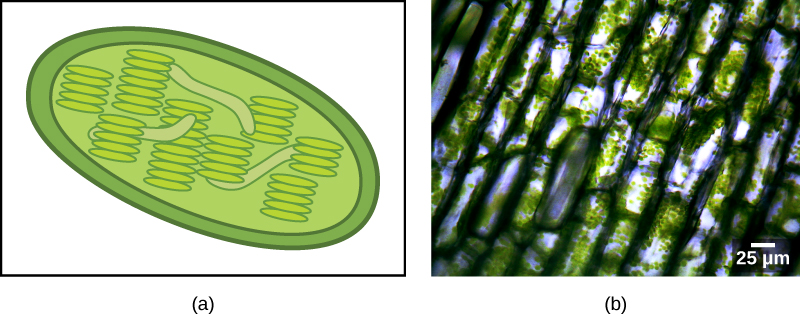
Like mitochondria, plastids appear to have an endosymbiotic origin. This hypothesis was also championed by Lynn Margulis. Plastids are derived from cyanobacteria that lived inside the cells of an ancestral, aerobic, heterotrophic eukaryote. This is called primary endosymbiosis, and plastids of primary origin are surrounded by two membranes. The best evidence is that this has happened twice in the history of eukaryotes. In one case, the common ancestor of the major lineage/supergroup Archaeplastida took on a cyanobacterial endosymbiont; in the other, the ancestor of the small amoeboid rhizarian taxon, Paulinella, took on a different cyanobacterial endosymbiont. Almost all photosynthetic eukaryotes are descended from the first event, and only a couple of species are derived from the other.
Cyanobacteria are a group of Gram-negative bacteria with all the conventional structures of the group. However, unlike most prokaryotes, they have extensive, internal membrane-bound sacs called thylakoids. Chlorophyll is a component of these membranes, as are many of the proteins of the light reactions of photosynthesis. Cyanobacteria also have the peptidoglycan wall and lipopolysaccharide layer associated with Gram-negative bacteria.
Chloroplasts of primary origin have thylakoids, a circular DNA chromosome, and ribosomes similar to those of cyanobacteria. Each chloroplast is surrounded by two membranes. In the group of Archaeplastida called the glaucophytes and in Paulinella, a thin peptidoglycan layer is present between the outer and inner plastid membranes. All other plastids lack this relictual cyanobacterial wall. The outer membrane surrounding the plastid is thought to be derived from the vacuole in the host, and the inner membrane is thought to be derived from the plasma membrane of the symbiont.
There is also, as with the case of mitochondria, strong evidence that many of the genes of the endosymbiont were transferred to the nucleus. Plastids, like mitochondria, cannot live independently outside the host. In addition, like mitochondria, plastids are derived from the division of other plastids and never built from scratch. Researchers have suggested that the endosymbiotic event that led to Archaeplastida occurred 1 to 1.5 billion years ago, at least 5 hundred million years after the fossil record suggests that eukaryotes were present.
Not all plastids in eukaryotes are derived directly from primary endosymbiosis. Some of the major groups of algae became photosynthetic by secondary endosymbiosis, that is, by taking in either green algae or red algae (both from Archaeplastida) as endosymbionts (Figure \(\PageIndex{3}\)). Numerous microscopic and genetic studies have supported this conclusion. Secondary plastids are surrounded by three or more membranes, and some secondary plastids even have clear remnants of the nucleus of endosymbiotic alga. Others have not “kept” any remnants. There are cases where tertiary or higher-order endosymbiotic events are the best explanations for plastids in some eukaryotes.
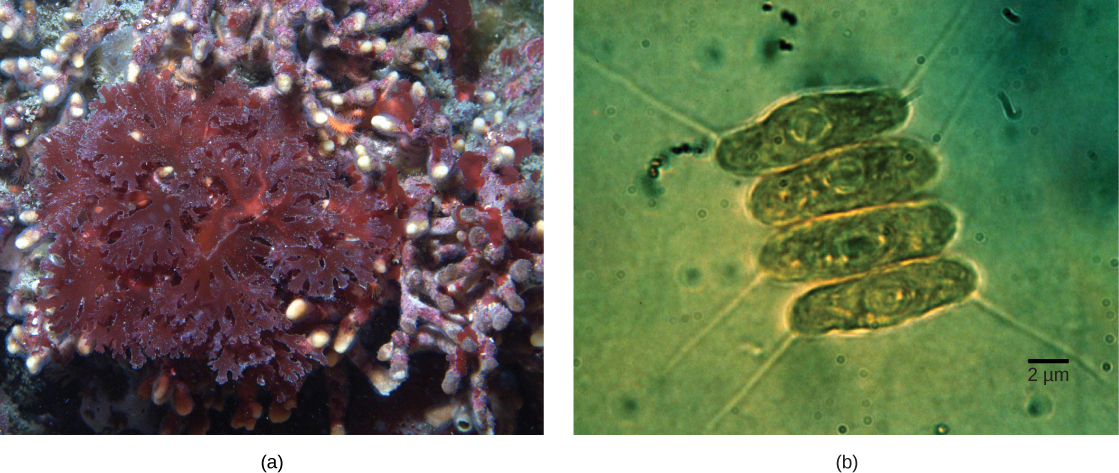
Art Connection

What evidence is there that mitochondria were incorporated into the ancestral eukaryotic cell before chloroplasts?
Evolution Connection: Secondary Endosymbiosis in Chlorarachniophytes
Endosymbiosis involves one cell engulfing another to produce, over time, a coevolved relationship in which neither cell could survive alone. The chloroplasts of red and green algae, for instance, are derived from the engulfment of a photosynthetic cyanobacterium by an early prokaryote.
This leads to the question of the possibility of a cell containing an endosymbiont to itself become engulfed, resulting in a secondary endosymbiosis. Molecular and morphological evidence suggest that the chlorarachniophyte protists are derived from a secondary endosymbiotic event. Chlorarachniophytes are rare algae indigenous to tropical seas and sand that can be classified into the rhizarian supergroup. Chlorarachniophytes extend thin cytoplasmic strands, interconnecting themselves with other chlorarachniophytes, in a cytoplasmic network. These protists are thought to have originated when a eukaryote engulfed a green alga, the latter of which had already established an endosymbiotic relationship with a photosynthetic cyanobacterium (Figure \(\PageIndex{5}\)).
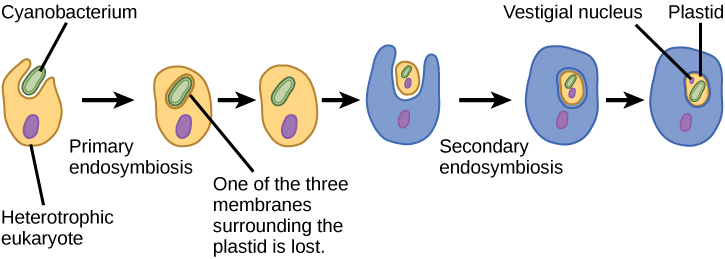
Several lines of evidence support that chlorarachniophytes evolved from secondary endosymbiosis. The chloroplasts contained within the green algal endosymbionts still are capable of photosynthesis, making chlorarachniophytes photosynthetic. The green algal endosymbiont also exhibits a stunted vestigial nucleus. In fact, it appears that chlorarachniophytes are the products of an evolutionarily recent secondary endosymbiotic event. The plastids of chlorarachniophytes are surrounded by four membranes: The first two correspond to the inner and outer membranes of the photosynthetic cyanobacterium, the third corresponds to the green alga, and the fourth corresponds to the vacuole that surrounded the green alga when it was engulfed by the chlorarachniophyte ancestor. In other lineages that involved secondary endosymbiosis, only three membranes can be identified around plastids. This is currently rectified as a sequential loss of a membrane during the course of evolution.
The process of secondary endosymbiosis is not unique to chlorarachniophytes. In fact, secondary endosymbiosis of green algae also led to euglenid protists, whereas secondary endosymbiosis of red algae led to the evolution of dinoflagellates, apicomplexans, and stramenopiles.
Summary
The oldest fossil evidence of eukaryotes is about 2 billion years old. Fossils older than this all appear to be prokaryotes. It is probable that today’s eukaryotes are descended from an ancestor that had a prokaryotic organization. The last common ancestor of today’s Eukarya had several characteristics, including cells with nuclei that divided mitotically and contained linear chromosomes where the DNA was associated with histones, a cytoskeleton and endomembrane system, and the ability to make cilia/flagella during at least part of its life cycle. It was aerobic because it had mitochondria that were the result of an aerobic alpha-proteobacterium that lived inside a host cell. Whether this host had a nucleus at the time of the initial symbiosis remains unknown. The last common ancestor may have had a cell wall for at least part of its life cycle, but more data are needed to confirm this hypothesis. Today’s eukaryotes are very diverse in their shapes, organization, life cycles, and number of cells per individual.
Art Connections
Figure \(\PageIndex{4}\): What evidence is there that mitochondria were incorporated into the ancestral eukaryotic cell before chloroplasts?
- Answer
-
All eukaryotic cells have mitochondria, but not all eukaryotic cells have chloroplasts.
Glossary
- endosymbiosis
- engulfment of one cell within another such that the engulfed cell survives, and both cells benefit; the process responsible for the evolution of mitochondria and chloroplasts in eukaryotes
- endosymbiotic theory
- theory that states that eukaryotes may have been a product of one cell engulfing another, one living within another, and evolving over time until the separate cells were no longer recognizable as such
- plastid
- one of a group of related organelles in plant cells that are involved in the storage of starches, fats, proteins, and pigments