
5.4: Photosynthesis
- Page ID
- 69383

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Explain the relevance of photosynthesis to other living things
- Describe the main structures involved in photosynthesis
- Identify the substrates and products of photosynthesis
- Summarize the process of photosynthesis
Main Structures and Summary of Photosynthesis
Photosynthesis is a multi-step process that requires sunlight, carbon dioxide (which is low in energy), and water as substrates (Figure \(\PageIndex{1}\)). After the process is complete, it releases oxygen and produces glyceraldehyde-3-phosphate (GA3P), simple carbohydrate molecules (which are high in energy) that can subsequently be converted into glucose, sucrose, or any of dozens of other sugar molecules. These sugar molecules contain energy and the energized carbon that all living things need to survive.

The following is the chemical equation for photosynthesis (Figure \(\PageIndex{2}\)):
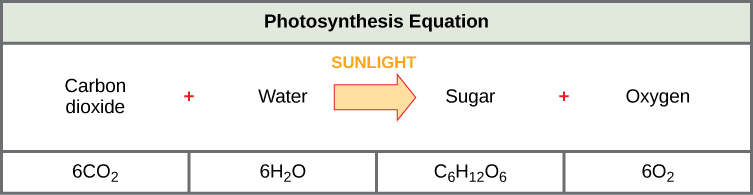
Although the equation looks simple, the many steps that take place during photosynthesis are actually quite complex. Before learning the details of how photoautotrophs turn sunlight into food, it is important to become familiar with the structures involved.
In plants, photosynthesis generally takes place in leaves, which consist of several layers of cells. The process of photosynthesis occurs in a middle layer called the mesophyll. The gas exchange of carbon dioxide and oxygen occurs through small, regulated openings called stomata (singular: stoma), which also play roles in the regulation of gas exchange and water balance. The stomata are typically located on the underside of the leaf, which helps to minimize water loss. Each stoma is flanked by guard cells that regulate the opening and closing of the stomata by swelling or shrinking in response to osmotic changes.
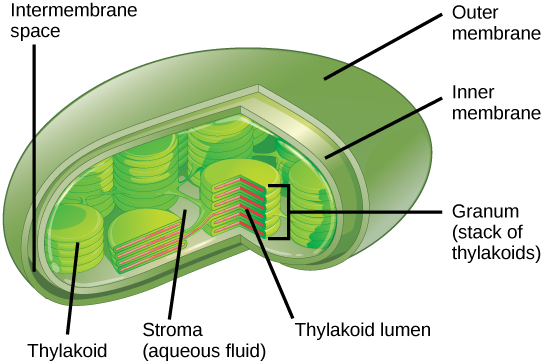
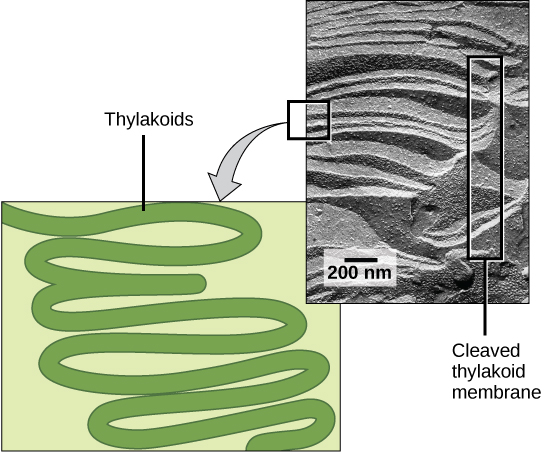
In all autotrophic eukaryotes, photosynthesis takes place inside an organelle called a chloroplast. For plants, chloroplast-containing cells exist in the mesophyll. Chloroplasts have a double membrane envelope (composed of an outer membrane and an inner membrane). Within the chloroplast are stacked, disc-shaped structures called thylakoids. Embedded in the thylakoid membrane is chlorophyll, a pigment (molecule that absorbs light) responsible for the initial interaction between light and plant material, and numerous proteins that make up the electron transport chain. The thylakoid membrane encloses an internal space called the thylakoid lumen. As shown in Figure \(\PageIndex{3}\), a stack of thylakoids is called a granum, and the liquid-filled space surrounding the granum is called stroma or “bed” (not to be confused with stoma or “mouth,” an opening on the leaf epidermis).
The Two Parts of Photosynthesis
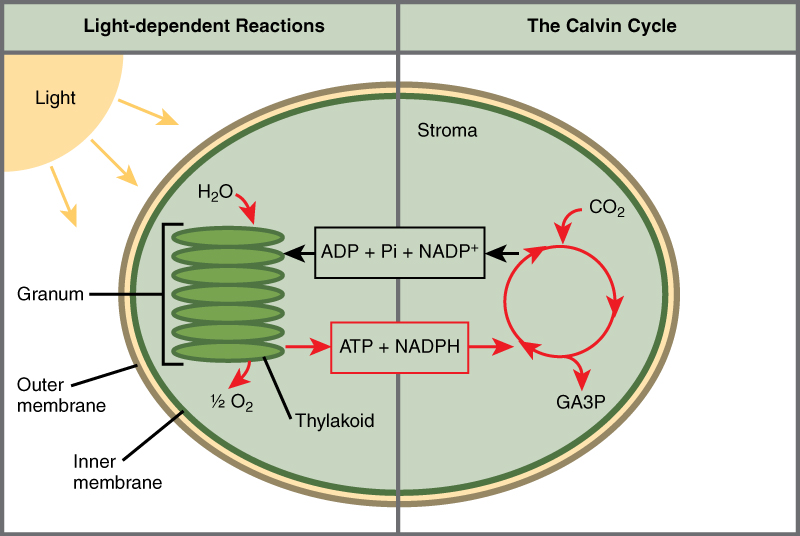
Photosynthesis takes place in two sequential stages: the light-dependent reactions and the light independent-reactions. In the light-dependent reactions, energy from sunlight is absorbed by chlorophyll and that energy is converted into stored chemical energy. In the light-independent reactions, the chemical energy harvested during the light-dependent reactions drive the assembly of sugar molecules from carbon dioxide. Therefore, although the light-independent reactions do not use light as a reactant, they require the products of the light-dependent reactions to function. In addition, several enzymes of the light-independent reactions are activated by light. The light-dependent reactions utilize certain molecules to temporarily store the energy: These are referred to as energy carriers. The energy carriers that move energy from light-dependent reactions to light-independent reactions can be thought of as “full” because they are rich in energy. After the energy is released, the “empty” energy carriers return to the light-dependent reaction to obtain more energy. Figure \(\PageIndex{4}\) illustrates the components inside the chloroplast where the light-dependent and light-independent reactions take place.
The Light-Dependent Reactions of Photosynthesis
Light energy initiates the process of photosynthesis when pigments absorb the light. Different kinds of organic pigments, whether in the human retina or the chloroplast thylakoid, have each evolved to absorb only certain wavelengths (colors) of light. Pigments reflect or transmit the wavelengths they cannot absorb, making them appear in the corresponding color. Plants pigment molecules absorb only light in the wavelength range of 700 nm to 400 nm; plant physiologists refer to this range for plants as photosynthetically active radiation.
Chlorophylls and carotenoids are the two major classes of photosynthetic pigments found in plants and algae; each class has multiple types of pigment molecules. There are five major chlorophylls: a, b, c and d and a related molecule found in prokaryotes called bacteriochlorophyll. Chlorophyll a and chlorophyll b are found in higher plant chloroplasts and will be the focus of the following discussion. With dozens of different forms, carotenoids are a much larger group of pigments. The carotenoids found in fruit—such as the red of tomato (lycopene), the yellow of corn seeds (zeaxanthin), or the orange of an orange peel (β-carotene)—are used as advertisements to attract seed dispersers. In photosynthesis, carotenoids function as photosynthetic pigments that are very efficient molecules for the disposal of excess energy. When a leaf is exposed to full sun, the light-dependent reactions are required to process an enormous amount of energy; if that energy is not handled properly, it can do significant damage. Therefore, many carotenoids reside in the thylakoid membrane, absorb excess energy, and safely dissipate that energy as heat.
Each type of pigment can be identified by the specific pattern of wavelengths it absorbs from visible light, which is the absorption spectrum. The graph in Figure \(\PageIndex{5}\) shows the absorption spectra for chlorophyll a, chlorophyll b, and a type of carotenoid pigment called β-carotene (which absorbs blue and green light). Notice how each pigment has a distinct set of peaks and troughs, revealing a highly specific pattern of absorption. Chlorophyll a absorbs wavelengths from either end of the visible spectrum (blue and red), but not green. Because green is reflected or transmitted, chlorophyll appears green. Carotenoids absorb in the short-wavelength blue region, and reflect the longer yellow, red, and orange wavelengths.

The overall function of light-dependent reactions is to convert solar energy into chemical energy in the form of NADPH and ATP. Protein complexes and pigment molecules work together to produce NADPH and ATP. The actual step that converts light energy into chemical energy takes place in a multiprotein complex called a photosystem, two types of which are found embedded in the thylakoid membrane, photosystem II (PSII) and photosystem I (PSI) (Figure \(\PageIndex{7}\)). The two complexes differ on the basis of what they oxidize (that is, the source of the low-energy electron supply) and what they reduce (the place to which they deliver their energized electrons). Both photosystems have the same basic structure; a number of antenna proteins to which the chlorophyll molecules are bound surround the reaction center where the photochemistry takes place. In short, the light energy has now been captured by biological molecules but is not stored in any useful form yet. The energy is transferred from chlorophyll to chlorophyll until eventually (after about a millionth of a second), it is delivered to the reaction center. Up to this point, only energy has been transferred between molecules, not electrons.

Figure \(\PageIndex{7}\): A photosystem consists of a light-harvesting complex and a reaction center. Pigments in the light-harvesting complex pass light energy to two special chlorophyll a molecules in the reaction center. The light excites an electron from the chlorophyll a pair, which passes to the primary electron acceptor. The excited electron must then be replaced. In the photosystem II (PSII) reaction center, energy from sunlight is used to extract electrons from water. The electrons travel through the chloroplast electron transport chain to photosystem I (PSI), which reduces NADP+ to NADPH. The electron transport chain moves protons across the thylakoid membrane into the lumen. At the same time, splitting of water adds protons to the lumen, and reduction of NADPH removes protons from the stroma. The net result is a low pH in the thylakoid lumen, and a high pH in the stroma. ATP synthase uses this electrochemical gradient to make ATP.
The reaction center in PSII contains a pair of chlorophyll a molecules with a special property. Those two chlorophylls can undergo oxidation upon excitation; they can actually give up an electron. It is at this step in photosynthesis, that light energy is converted into an excited electron. All of the subsequent steps involve getting that electron onto the energy carrier NADPH for delivery to the Calvin cycle where the electron is deposited onto carbon for long-term storage in the form of a carbohydrate. The reaction center of PSII extracts low-energy electrons from water, splitting two molecules of water to release four electrons, four hydrogen atoms, and one molecule of diatomic O2 gas. The reaction center of PSII then delivers its high-energy electrons, one at the time, to the primary electron acceptor, and through the electron transport chain.
The cytochrome complex, an enzyme composed of two protein complexes, transfers the electrons from the carrier molecule plastoquinone (Pq) to the protein plastocyanin (Pc), thus enabling both the transfer of protons across the thylakoid membrane and the transfer of electrons from PSII to PSI (Figure \(\PageIndex{7}\)). As electrons move through the proteins that reside between PSII and PSI, they lose energy. That energy is used to move hydrogen atoms from the stromal side of the membrane to the thylakoid lumen. Those hydrogen atoms, plus the ones produced by splitting water, accumulate in the thylakoid lumen and create a concentration gradient. The passive diffusion of hydrogen ions from high concentration (in the thylakoid lumen) to low concentration (in the stroma) through a specialized protein channel called the ATP synthase. The energy released by the hydrogen ion stream as it passes through the thylakoid membrane allows ATP synthase to attach a third phosphate group to ADP, which forms a molecule of ATP (Figure \(\PageIndex{7}\)).
Because the electrons have lost energy prior to their arrival at PSI, they must be re-energized by PSI, hence, another photon is absorbed by the PSI antenna. That energy is relayed to the PSI reaction center which is oxidized and sends a high-energy electron to NADP+ to form NADPH. Thus, PSII captures the energy to create proton gradients to make ATP, and PSI captures the energy to reduce NADP+ into NADPH.
The Light-Independent Reactions of Photosynthesis
After the energy from the sun is converted into chemical energy temporarily stored in the bonds of ATP and NADPH molecules, the cell has the fuel needed to build carbohydrate molecules for long-term energy storage. The products of the light-dependent reactions, ATP and NADPH, have lifespans in the range of millionths of seconds, whereas the products of the light-independent reactions (carbohydrates and other forms of reduced carbon) can survive for hundreds of millions of years. In plants, carbon dioxide (CO2) enters the leaves through stomata, where it diffuses over short distances through intercellular spaces until it reaches the mesophyll cells. Once in the mesophyll cells, CO2 diffuses into the stroma of the chloroplast—the site of light-independent reactions of photosynthesis (Figure \(\PageIndex{8}\)). The light-independent reactions (also known as the Calvin cycle) can be organized into three basic stages: fixation, reduction, and regeneration.
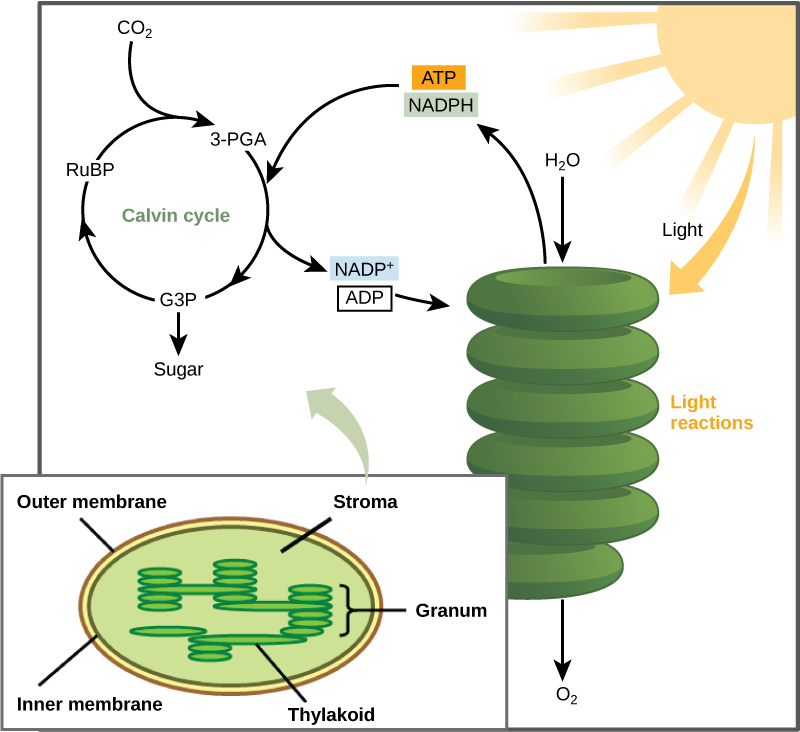
Stage 1: Fixation:
In the stroma, in addition to CO2, two other components are present to initiate the light-independent reactions: an enzyme called ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO), and three molecules of ribulose-1,5-bisphosphate (RuBP), as shown in Figure 5.4.1.25.4.1.2. RuBP has five atoms of carbon, flanked by two phosphates.
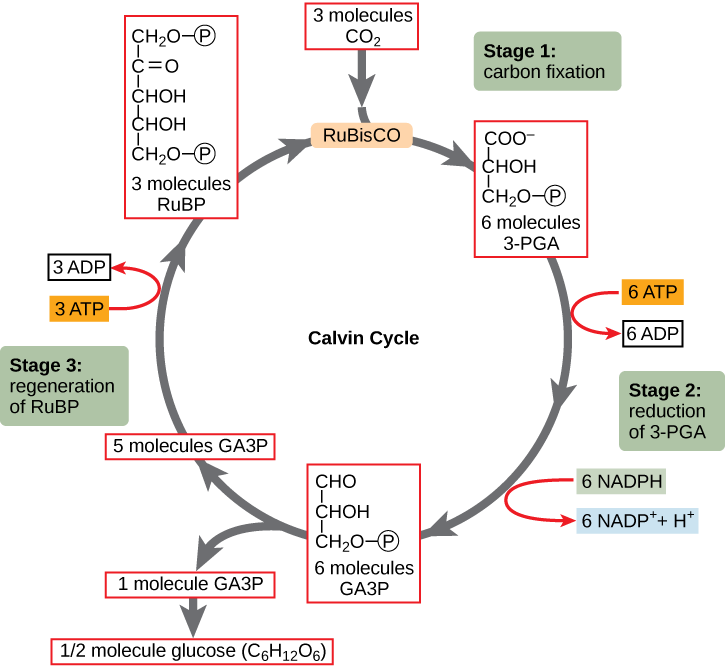
RuBisCO catalyzes a reaction between CO2 and RuBP. For each CO2 molecule that reacts with one RuBP, two molecules of another compound, 3-phosphoglycerate (3-PGA), form. 3-PGA has three carbon atoms and one phosphate. Each turn of the cycle involves only one RuBP and one carbon dioxide and forms two molecules of 3-PGA. The number of carbon atoms remains the same, as the atoms move to form new bonds during the reactions (3 atoms from 3 CO2 + 15 atoms from 3 RuBP = 18 atoms in 3 atoms of 3-PGA). This process is called carbon fixation, because CO2 is “fixed” from an inorganic form into organic molecules.
Stage 2: Reduction:
ATP and NADPH are used to convert the six molecules of 3-PGA into six molecules of a chemical called glyceraldehyde 3-phosphate (G3P). That is a reduction reaction because it involves the gain of electrons by 3-PGA. Recall that reduction is the gain of an electron by an atom or molecule. Six molecules of both ATP and NADPH are used. For ATP, energy is released with the loss of the terminal phosphate atom, converting it into ADP; for NADPH, both energy and a hydrogen atom are lost, converting it into NADP+. Both of these molecules return to the nearby light-dependent reactions to be reused and re-energized.
Stage 3: Regeneration:
Interestingly, at this point, only one of the G3P molecules leaves the light-independent reactions and is sent to the cytoplasm to contribute to the formation of other compounds needed by the plant. Because the G3P exported from the chloroplast has three carbon atoms, it takes three “turns” of the cycle to fix enough net carbon to export one G3P. But each turn makes two G3P, thus three turns make six G3P. One of these six is exported while the remaining five G3P molecules remain in the cycle and are used to regenerate RuBP, which enables the system to prepare for more CO2 to be fixed. Three more molecules of ATP are used in these regeneration reactions.
Because the carbohydrate molecule has six carbon atoms, it takes six turns of the Calvin cycle to make one carbohydrate molecule (one for each carbon dioxide molecule fixed). The remaining G3P molecules regenerate RuBP, which enables the system to prepare for the carbon-fixation step. ATP is also used in the regeneration of RuBP. These six turns require energy input from at total of 12 ATP molecules and 12 NADPH molecules in the reduction step and a total of 6 ATP molecules in the regeneration step.
Photosynthesis in Prokaryotes
The two parts of photosynthesis—the light-dependent reactions and the Calvin cycle—have been described, as they take place in chloroplasts. However, prokaryotes, such as cyanobacteria, lack membrane-bound organelles. Prokaryotic photosynthetic autotrophic organisms have infoldings of the plasma membrane for chlorophyll attachment and photosynthesis (Figure \(\PageIndex{10}\)). It is here that organisms like cyanobacteria can carry out photosynthesis.
Photorespiration and C4 and CAM Plants
Different plant species have adaptations that allow them to do different variations of the light-independent reactions. These are called photosynthetic pathways. Plants are classified as C3, C4, or CAM depending on their use of these pathways, but note that some plants can switch photosynthetic pathways depending on environmental conditions. The process for light-independent reactions described in the previous section was the C3 pathway: the compound formed during fixation (3-PGA) has three carbon atoms.
Photosynthesis in dry-climate plants (Figure \(\PageIndex{11}\)) has evolved with adaptations that conserve water. In the harsh dry heat, every drop of water and precious energy must be used to survive. Two adaptations have evolved in such plants. In one form, a more efficient use of CO2 allows plants to photosynthesize even when CO2 is in short supply, as when the stomata are closed on hot days. The other adaptation performs preliminary reactions of the Calvin cycle at night, because opening the stomata at this time conserves water due to cooler temperatures. In addition, this adaptation has allowed plants to carry out low levels of photosynthesis without opening stomata at all, an extreme mechanism to face extremely dry periods.

Figure \(\PageIndex{11}\): Living in the harsh conditions of the desert has led plants like this cactus to evolve variations in reactions outside the Calvin cycle. These variations increase efficiency and help conserve water and energy. (credit: Piotr Wojtkowski)
As its name suggests, ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO) catalyzes two different reactions. The first is adding CO2 to ribulose-1,5- bisphosphate (RuBP) — the carboxylase activity. The second is adding O2 to RuBP — the oxygenase activity. The oxygenase activity of RuBisCO forms the three-carbon molecule 3-phosphoglycerate (3-PGA), just as in the light-independent reactions, and the two-carbon molecule glycolate. The glycolate enters peroxisomes, where it uses O2 to form intermediates that enter mitochondria where they are broken down to CO2. So this process uses O2 and liberates CO2 as aerobic cellular respiration does, which is why it is called photorespiration. It undoes the work of photosynthesis, which is to build sugars. Which action of RuBisCO predominates depends on the relative concentrations of O2 and CO2 with high CO2, low O2 favoring the carboxylase action and high O2, low CO2 favoring the oxygenase action. The light reactions of photosynthesis liberate oxygen, and more oxygen dissolves in the cytosol of the cell at higher temperatures. Therefore, high light intensities and high temperatures (above ~ 30°C) favor the second reaction and result in photorespiration.
One solution to photorespiration is for plants to open their stomata to release O2 and obtain CO2. However, if conditions are hot or dry, this will result in too much water loss (transpiration). For this reason, C3 plants, which only do the C3 pathway and do not use the C4 pathway to prevent photorespiration (see below), do best in cool, moist areas. Rice and potatoes are examples of C3 plants.
C4 Plants
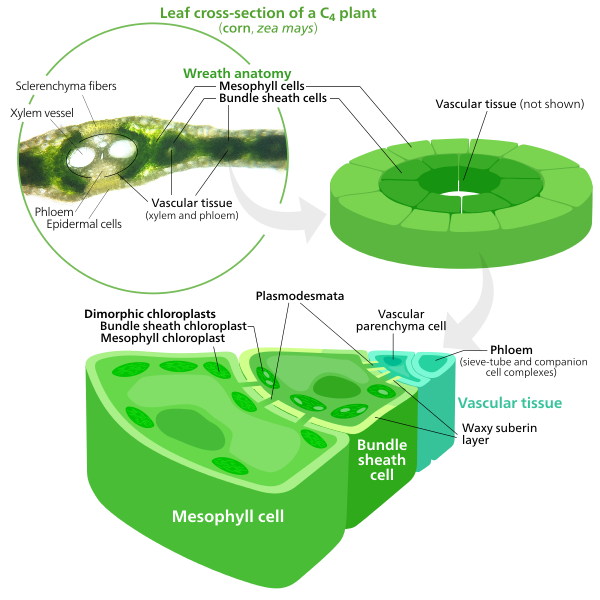
Many angiosperms have developed adaptations which minimize the losses to photorespiration. They all use a supplementary method of CO2 uptake which initially forms a four-carbon molecule compared to the two three-carbon molecules that are initially formed in the C3 pathway. Hence, these plants are called C4 plants. Note that C4 plants will eventually conduct the light-independent reactions (C3 pathway), but they form a four-carbon molecule first. C4 plants have structural changes in their leaf anatomy so that synthesizing the four-carbon sugar (the C4 pathway) and resuming the light-independent reactions (C3 pathways) are separated in different parts of the leaf with RuBisCO sequestered where the CO2 level is high and the O2 level low. After entering through stomata, CO2 diffuses into a mesophyll cell (Figure \(\PageIndex{12}\)). Being close to the leaf surface, these cells are exposed to high levels of O2, but they have no RuBisCO so cannot start photorespiration (nor the light-independent reactions).
Instead, the CO2 is inserted into a three-carbon compound called phosphoenolpyruvic acid (PEP) forming the four-carbon compound oxaloacetic acid. Oxaloacetic acid is converted into malic acid or aspartic acid (both have 4 carbons), which is transported through plasmodesmata into a bundle sheath cell. Bundle sheath cells are deep in the leaf, so atmospheric oxygen cannot diffuse easily to them (Figure \(\PageIndex{13}\)). Additionally, they often have thylakoids with reduced photosystem II complexes (the one that produces O2). Both of these features keep oxygen levels low in bundle sheath cells, which is where the four-carbon compound is broken down into carbon dioxide, which enters the light-independent reactions (C3 pathway) to form sugars and pyruvic acid, which is transported back to a mesophyll cell where it is converted back into PEP.
These C4 plants are well adapted to (and likely to be found in) habitats with high daytime temperatures and intense sunlight. Because they use the C4 pathway to prevent photorespiration, they do not have to open their stomata to the same extent as C3 plants and can thus conserve water. Some examples crabgrass, corn (maize), sugarcane, and sorghum. Although comprising only ~3% of the angiosperms by species, C4 plants are responsible for ~25% of all the photosynthesis on land.
CAM plants
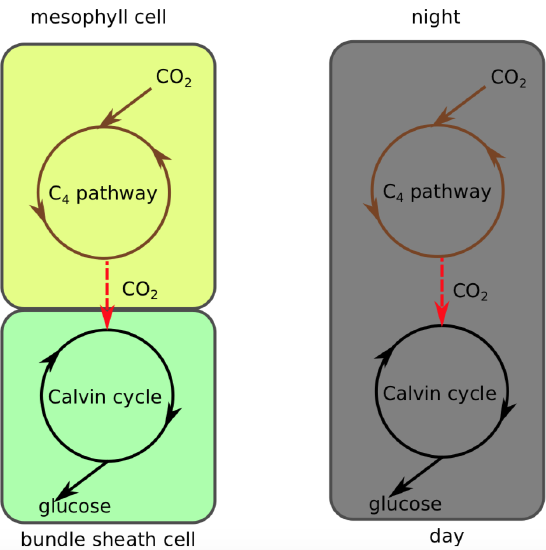
CAM stands for crassulacean acid metabolism because it was first studied in members of the plant family Crassulaceae. CAM plants also do the C4 pathway. However, instead of segregating the C4 and C3 pathways in different parts of the leaf, CAM plants separate them in time instead (Table \(\PageIndex{1}\)). As a result, CAM plants do not need to open their stomata in the daytime to reduce photorespiration because they have already formed a four-carbon molecule at night that can be broken down to release carbon dioxide during the day.
| Night | Morning |
|---|---|
|
|
CAM plants thus thrive in conditions of high daytime temperatures, intense sunlight, and low soil moisture. Some examples of CAM plants include cacti (Figure \(\PageIndex{14}\)), pineapples, all epiphytic bromeliads, sedums, and the "ice plant" that invade the California coast line.

Further "Reading"
For more on the Calvin Cycle see this Calvin Cycle Video from TEDEd or this video Phyotosynthesis Animation from HHMI Biointeractive.
Contributors and Attributions
Modified by Kyle Whittinghill (University of Pittsburgh) and Melissa Ha from
Connie Rye (East Mississippi Community College), Robert Wise (University of Wisconsin, Oshkosh), Vladimir Jurukovski (Suffolk County Community College), Jean DeSaix (University of North Carolina at Chapel Hill), Jung Choi (Georgia Institute of Technology), Yael Avissar (Rhode Island College) among other contributing authors. Original content by OpenStax (CC BY 4.0; Download for free at http://cnx.org/contents/185cbf87-c72...f21b5eabd@9.87).
- 8.3 Using Light Energy to Make Organic Molecules from Biology 2e by OpenStax (licensed CC-BY). Access for free at openstax.org.
- 3.3 Enzymatic Stage from Introduction to Botany by Alexey Shipunov (public domain)
- 13.6: Light-independent Reactions by Melissa Ha, Maria Morrow, & Kammy Algiers, is licensed CC BY-SA
- 8.1: Overview of Photosynthesis by OpenStax, is licensed CC BY
- 8.2: The Light-Dependent Reactions of Photosynthesis by OpenStax, is licensed CC BY
- 5.3: The Calvin Cycle by OpenStax, is licensed CC BY
- 13.7: Photorespiration and Photosynthetic Pathways by Melissa Ha, Maria Morrow, & Kammy Algiers, is licensed CC BY-SA
- 16.2E Photorespiration and C4 Plants from Biology by John W. Kimball (licensed CC-BY)
- Essentials of Environmental Science by Kamala Doršner is licensed under CC BY 4.0.