
Gluconeogenesis
- Page ID
- 1334
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Gluconeogenesis is the synthesis of glucose. It is basically glycolysis run backwards; three new reactions (involving four new enzymes) make the standard free energy favorable.
- Glycolysis: \( \Delta{G}_o’ = -74 \, \text{kcal/mol} \)
- Gluconeogenesis: \( \Delta{G}_o' = -36 \, \text{kcal/mol} \)
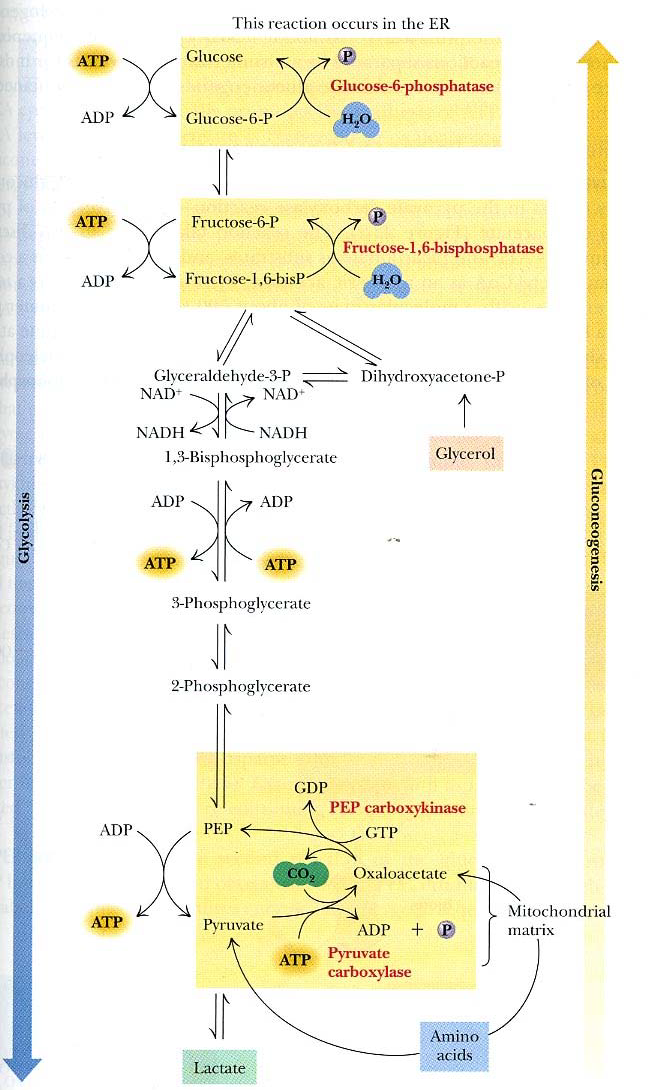
For every molecule of glucose synthesized from two molecules of pyruvate, 4 ATP, 2 GTP, and 2 NADH are used.
In the Mitochondria
Pyruvate + ATP \( \rightarrow \) Oxaloacetate + ADP + P
Oxaloacetate + NADH \( \rightarrow \) Malate + NAD+
The conversion to malate allows the molecule to be transported out of the mitochondria. Once in the cytoplasm, it is converted back to oxaloacetate.
In the Cytoplasm
Malate + NAD+ \( \rightarrow \) Oxaloacetate + NADH
Oxaloacetate + GTP \( \rightarrow \) PEP + GDP
From here, it goes through the same intermediates as glycolysis. The last reaction happens in the endoplasmic reticulum.
In the Endoplasmic Reticulum
G6P \( \rightarrow \) glucose (catalyst: glucose-6-phosphatase)
A glucose transporter shuttles the glucose out into the extracellular space.
Regulation
| Regulated Reactions | Glycolysis | Gluconeogenesis |
| Glucose \( \rightleftharpoons \) G6P | Hexokinase: G6P (-) | Glucose-6-phosphatase: [G6P] (substrate level control) |
| F6P \( \rightleftharpoons \) F1,6BP | Phosphofructokinase: F2,6BP (+); AMP (+); ATP (-); citrate (-) | Fructose-1,6-bisphosphatase: F26BP (-); AMP (-) |
| PEP \( \rightleftharpoons \) Pyruvate | Pyruvate kinase: F1,6BP (+); acetyl CoA (-); ATP (-); alanine (-); cAMP-dependent phosphorylation (-) | Pyruvate carboxylase: Acetyl-CoA (+) |
These reactions are tightly controlled so that glycolysis and gluconeogenesis are not run at the same time. If they were, the F1,6BP \( \rightleftharpoons \) F6P reaction could turn into a futile cycle, using up ATP without progressing in either direction.
Glyoxylate Cycle
Plants and bacteria can convert acetyl-CoA to glucose via the glyoxylate cycle. It is a modified version of the TCA cycle; an extra malate is produced which can be converted to glucose. Since animals lack this cycle, they cannot use acetyl-CoA to make glucose because it would stop the TCA cycle.
Starch/Glycogen Synthesis
Glucose is added to chains of glycogen for storage via starch/glycogen synthesis. Glucose is converted to G-6-P, then G-1-P. This is added to UDP, which gives glucose the free energy needed to add to the glycogen. (Plants use ADPG and ATP instead of UDPG and GTP.)
Glucose + ATP \( \rightarrow \) G-6-P + ATP
(catalyst: hexokinase)
G-6-P \( \rightarrow \) G-1-P
(catalyst: P-glucomutase)
UTP + G-1-P \( \rightarrow \) UDPG + PPi
(catalyst: UDPG pyrophosphorylase)
PPi + H2O \( \rightarrow \) 2Pi
The glucose of the UDP-glucose is added to the glycogen chain, leaving UDP.
Pentose Phosphate Pathway (PPP)
The PPP is a source of NADPH, which can be used in reductive anabolic pathways. The PPP can also produce ribose.