
5.1: Changes in Chromosome Number
- Page ID
- 25739

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Learning Objectives
- Describe nondisjunction and its outcomes.
- Distinguish between aneuploidy (example: nullisomy or trisomy) and polyploidy.
- Give an example of a human trisomy and explain why trisomies of some chromosomes are not tolerated (viable in offspring).
- Explain why organisms with an odd number of chromosome sets are usually sterile or not even viable.
- Recall that polyploidy is common in plants but rare in animals. Give an example of an advantage of polyploidy.
Single Chromosomes can be Lost or Gained by Nondisjunction
If something goes wrong during cell division, an entire chromosome may be lost and the cell will lack all of these genes. The causes behind these chromosome abnormalites and the consequences they have for the cell and the organism is the subject of this section.
Cause: Nondisjunction During Mitosis or Meiosis
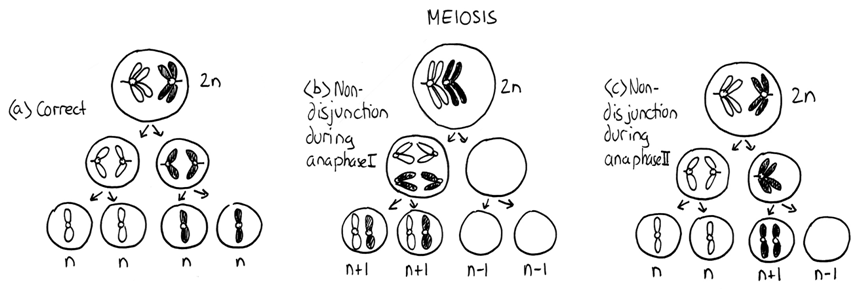
Segregation occurs in anaphase. In mitosis and meiosis II, sister chromatids (of replicated chromosomes) are normally pulled to opposite ends of the cell. In Meiosis I, it is homologous chromosomes, which are synapsed at that time, that segregate and move apart.
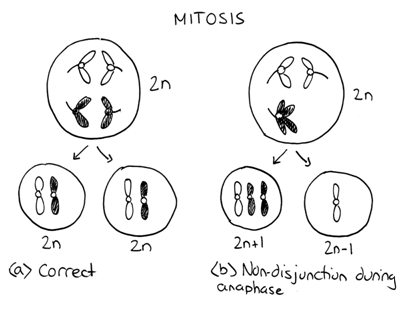
In rare cases, the sister chromatids (or paired chromosomes in Meiosis I) fail to separate. This failure to segregate properly is called nondisjunction and it can happen during mitosis, meiosis I, or meiosis II. This nondisjunction results in both chromatids (or chromosomes) moving to one pole and none at the other. One cell will have an extra copy and the other will lack a copy. Thus failure to segregate properly leads to unbalanced products.
Consequence: Decreased Viability
The result of a non-disjunction event is daughter cells that have an abnormal number of chromosomes. Cells, such as the parent cell in Figure 5.1.1, which have the proper number of chromosomes, are said to be euploid. The daughter cells have one too many or one too few chromosomes and are thus aneuploid. Even though both product cells have at least one copy of all genes, both cells will probably die. The reason is due to the loss or gain of a large number of genes. Genes produce an standard amount of product - either functional RNAs or proteins. The parent cell shown has a balanced genotype because it has two copies of all of its genes. Each of its genes produces half of the products needed by the cell. But if one of these cells suddenly had only one copy (or three copies) of an important gene, the amount of product would be either 50% (or 150%) of what was required. Such a change for a single gene could probably be tolerated by the cell and it would probably survive. But the sudden change to one copy (or three copies) of the hundreds or thousands of genes on an entire chromosome the results would be more than tolerable and be catastrophic for the daughter cells. They have what’s called an unbalanced genotype, which usually decreases their viability.
If a nondisjunction event occurs during either meiosis I or meiosis II, the result is an unbalanced gamete. The gamete will often be functional, but after fertilization the embryo will be genetically unbalanced. This usually leads to the death of the embryo. There are some exceptions to this in humans and these will be presented later in this chapter.
Changes in sets of chromosomes
Humans, like most animals and most eukaryotic genetic model organisms, are diploids because they have two copies of each autosome. This means that most of their cells have two homologous copies of each chromosome. In contrast, many plant species and even a few animal species are polyploids. This means they have more than two chromosome sets, and so have more than two homologs of each chromosome in each cell.
When the nuclear content changes by a whole chromosome set we call it a change in ploidy. Gametes are haploid (n) and thus most animals are diploid (2n), formed by the fusion of two haploid gametes. However, some species can exist as monoploid (1x), triploid (3x), tetraploid (4x), pentaploid (5x), hexaploid (6x), or higher.
Notation of ploidy
As we have already seen, the letter n is the number of chromosomes in a gamete, and 2n is the number of chromosomes following fertilization.
To distinguish polyploids, the letter x is used to represent chromosome sets. For example:
- For a tetraploid 2n=4x and a gamete has n=2x chromosomes.
- For a hexaploid 2n=6x and a gamete has n=3x chromosomes.
Query \(\PageIndex{1}\)
Male Bees are Monoploid
Monoploids, with only one set of chromosomes, are usually inviable in most species; however, in many species of Hymenoptera (bees, wasps, ants) the males are monoploid and develop from unfertilized eggs. These males don’t undergo meiosis for gametes; mitosis produces sperm. Females are diploid (from fertilized eggs) and produce eggs via meiosis. Female bees are diploid (2n=32) and are formed when an egg (n=16) is fertilized by a sperm (n=16). If an egg is not fertilized, it can still develop and the result is a n=16 male drone. Males are described as haploid (because they have the same number of chromosomes as a gamete) or monoploid (because they have only one chromosome set). Females produce eggs by meiosis while males produce sperm by mitosis. This form of sex determination produces more females, workers, which do the work than males, who are only needed for reproduction.

Polyploids can be stable or sterile
Like diploids (2n=2x), stable polyploids generally have an even number of copies of each chromosome: tetraploid (2n=4x), hexaploid (2n=6x), and so on. The reason for this is clear from a consideration of meiosis. Remembering that the purpose of meiosis is to reduce the sum of the genetic material by half, meiosis can equally divide an even number of chromosome sets, but not an odd number. Thus, polyploids with an odd number of chromosomes (e.g. triploids, 2n=3x) tend to be sterile, even if they are otherwise healthy.
The mechanism of meiosis in stable polyploids is essentially the same as in diploids: during metaphase I, homologous chromosomes pair with each other. Depending on the species, all of the homologs may be aligned together at metaphase, or in multiple separate pairs. For example, in a tetraploid, some species may form tetravalents in which the four homologs from each chromosome align together, or alternatively, two pairs of homologs may form two bivalents. Note that because that mitosis does not involve any pairing of homologous chromosomes, mitosis is equally effective in diploids, even-number polyploids, and odd-number polyploids.
Many Crop Plants are Hexaploid or Octoploid
Polyploid plants tend to be larger and healthier than their diploid counterparts. The strawberries sold in grocery stores come from octoploid (8x) strains and are much larger than the strawberries formed by wild diploid strains.
Bread wheat is a hexaploid (2n=6x) strain. This species is derived from the combination of three other wheat species, T. monococcum, T. searsii, and T. tauschii. Each of these chromosome sets has 7 chromosomes so the diploid species are 2n=2x=14 and bread wheat is 2n=6x=42 and has 14 chromosomes from each species. Bread wheat is viable because each chromosome behaves independently during mitosis. The species is also fertile because during meiosis I the equivalent chromosomes from each species can pair each other during meiosis I. Thus, even in a polyploid, homologous chromosomes can segregate equally and gene balance can be maintained.
Bananas, Watermelons, and Other Seedless Plants are Triploid
The bananas found in grocery stores are a seedless variety called Cavendish. They are a triploid variety (chromosome sets = AAA) of a normally diploid species called Musa acuminata (AA). Cavendish plants are viable because mitosis can occur. However they are sterile because the chromosomes cannot pair properly during meiosis I. During prophase I there are three copies of each chromosome trying to “pair” with each other. Because proper chromosome segregation in meiosis fails, seeds cannot be made and the result is a fruit that is easier to eat because there are no seeds to spit out.
Why are triploids sterile?
Remember that during meiosis in a diploid there are four sister chromatids for each chromosome and 4 cells (gametes) produced. Each gamete inherits one copy of each chromosome. In a triploid after DNA replication (S phase), there will be six sister chromatids for each chromosome, but meiosis I and II still undergo the same process.
- How many chromatids will be in each gamete?
- Will the number of copies of each chromosome be the same in a single gamete?
For a cell with three types of chromosomes (n=3), draw the chromosomes in a triploid cell. Show how those chromosomes might segregate during meiosis. What possibilities exist for number of chromosomes in each gamete?
Exercise \(\PageIndex{1}\)
Seedless watermelons are another common triploid. The haploid number (n) of standard watermelons is 11. How many chromosomes are present in a single triploid watermelon cell? The thin white seeds in these triploids are the result of unbalanced gametes, but rare black viable seeds are sometimes found in a seedless watermelon. What would have to happen for these viable seeds to be produced?

- Answer
-
A triploid watermelon has 3 sets of 11 chromosomes: 3 x 11 = 33.
Although most gametes will have unbalanced chromosomes (example: one copy of five chromosomes but two copies of six chromosomes), rare balanced gametes can be produced. For example, by chance a balanced gamete with one copy of all eleven chromosomes is fertilized by another balanced gamete with two copies of all chromosomes.
If triploids cannot make seeds, how do we obtain enough triploid individuals for cultivation? The answer depends on the plant species involved. In some cases, such as banana, it is possible to propagate the plant asexually; new progeny can simply be grown from cuttings from a triploid plant. On the other hand, seeds for seedless watermelon are produced sexually: a tetraploid watermelon plant is crossed with a diploid watermelon plant. Both the tetraploid and the diploid are fully fertile, and produce gametes with two (1n=2x) or one (1n=1x) sets of chromosomes, respectively. These gametes fuse to produce a zygote (2n=3x) that is able to develop normally into an adult plant through multiple rounds of mitosis, but is unable to compete normal meiosis or produce seeds.
Polyploids are often larger in size than their diploid relatives. Food plant cultivation takes advantage of this property. For example, most strawberries you eat are not diploid, but octoploid (8x).
Polyploidy in Animals
Polyploidy in animals is rare, essentially limited to lower forms, although some vertebrates (fish) have been identified as triploid. Cultivated oysters are an example of a viable and commercially relevant triploid animal. In the 1990s, biologists explored ways to alter the number of chromosome sets in the oyster Crassostrea gigas (Guo et al, 1992).
Triploid oysters
Triploid oysters are cultivated for human consumption (https://www.theatlantic.com/technology/archive/2014/09/todays-oysters-are-mutants/380858/). What are the advantages of triploidy in oysters? How are triploid oysters similar to triploid plants? Based on the observation of rare fertilized seeds in seedless watermelons, would you expect to find rare fertile triploid oysters?
Triploidy in humans is not a viable condition, occurring in as much as 20% of miscarriages that show chromosomal abnormalities (reviewed in Kolarski et al, 2017). Triploidy in humans may be caused by abnormal meiosis that produces a gamete with two sets of chromosomes or by fertilization of a single egg by two sperm.
References
Guo, Ximing, et al. “Genetic Consequences of Blocking Polar Body I with Cytochalasin B in Fertilized Eggs of the Pacific Oyster, Crassostrea Gigas: I. Ploidy of Resultant Embryos.” Biological Bulletin, vol. 183, no. 3, 1992, pp. 381–386. JSTOR, www.jstor.org/stable/1542013. Accessed 7 July 2020.
Kolarski M, Ahmetovic B, Beres M, et al. Genetic Counseling and Prenatal Diagnosis of Triploidy During the Second Trimester of Pregnancy. Med Arch. 2017;71(2):144-147. doi:10.5455/medarh.2017.71.144-147
Contributors and Attributions
Dr. Todd Nickle and Isabelle Barrette-Ng (Mount Royal University) The content on this page is licensed under CC SA 3.0 licensing guidelines.
Dr. Todd Nickle and Isabelle Barrette-Ng (Mount Royal University) The content on this page is licensed under CC SA 3.0 licensing guidelines.