4.5.1: Linkage and Mapping
- Page ID
- 32591

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Learning Objectives
- Understand that linked genes do not exhibit independent assortment because recombination will not always occur between the loci.
- Distinguish between parental and recombinant chromosomes, gametes, and offspring, and identify them in crosses.
- Calculate the map distance between loci given the phenotypes of offspring or predict phenotypes of offspring given the recombination frequency between loci. Use the distance to construct genetic maps based on data from two-point or three-point testcrosses.
Loci are locations of genes on chromosomes
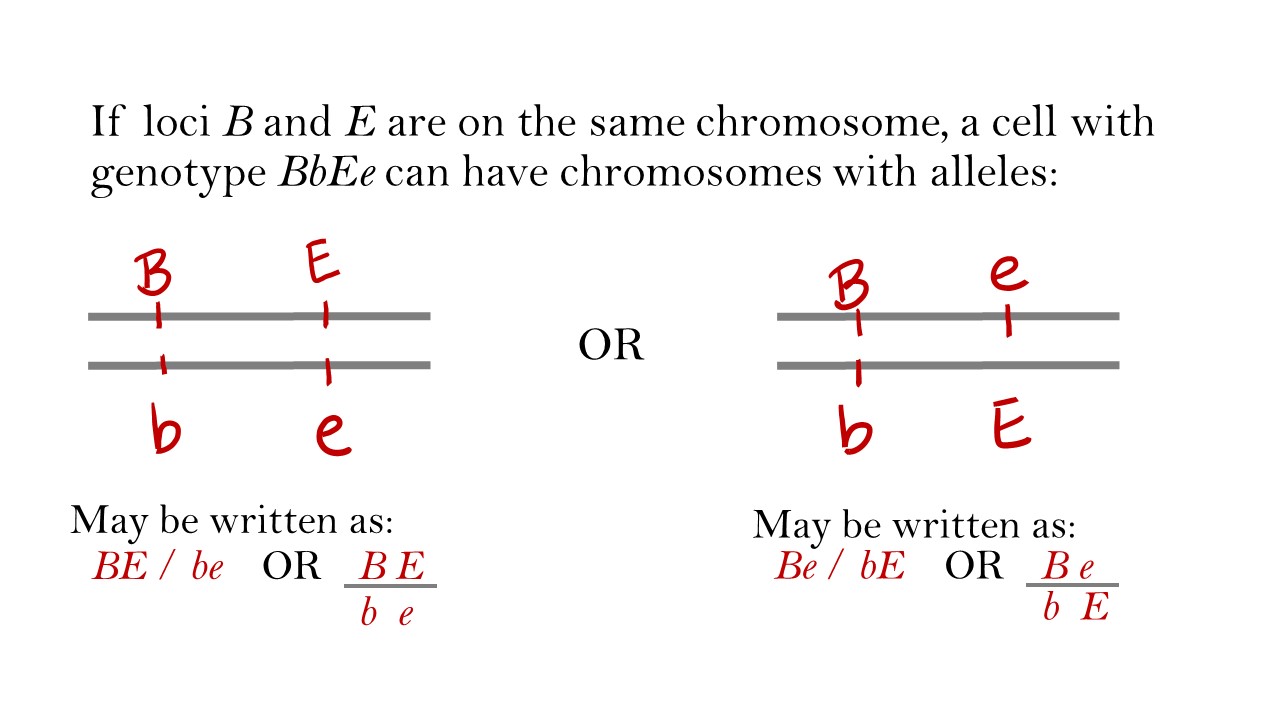
In many genetic crosses involving one or two genes, the gene can be representing by a name or a letter. However, when considering linked genes the location of each gene and allele often needs to be represented. For example, a dihybrid BbEe can have one chromosome with both dominant alleles (BE) or one chromosome with a dominant allele for one gene and recessive allele for the other (Be for example). The locations of the genes on the chromosomes are loci. For this section, remember that is the distance between loci that influences how often homologous recombination occurs between them between meiosis.
Exercise \(\PageIndex{1}\)
In Figure \(\PageIndex{1}\), why is a chromosome with Bb or Ee not shown as a possible arrangement?
- Answer
-
The loci are the locations of the genes on chromosomes. The B gene must be on the same position on each homologous chromosome in pair. The B gene is not located at two positions on one chromosome.
Effect of recombination on gamete possibilities
If loci B and E in the above example (Figure \(\PageIndex{1}\)) were on two different chromosomes, we would expect to obtain four gamete genotypes (25% each): BE, Be, bE, and be, as observed by independent assortment. However, if B and E in the above example were so close that homologous recombination (crossing over) never occurs between them during meiosis, then all types of gametes will not be observed. An organism with chromosomes BE / be could produce only gametes BE and be (50% each). An organism with chromosomes Be / bE could produce only gametes Be and bE (50% each).
Homologous recombination during meiosis I breaks and rejoins pieces of homologous chromosomes. If homologous recombination occurs between B and E then all four gametes will be possible. But at what frequency will each gamete be observed? The answer depends on how far apart they are! Recombination frequency is the percent of meioses in which homologous recombination exchanges two loci. In genetic mapping, this number expresses distance in map units (m.u.) or centiMorgans (cM) (named after geneticist Thomas Hunt Morgan).
How do geneticists know if recombination has occurred? Use a testcross. As shown in the next video, the map distance between loci B and E is determined by the number of recombinant offspring.
Remember:
- The # of recombinant offspring / total # of offspring x 100% = recombination frequency
- Recombination frequency = map units = centiMorgan (cM)
Example \(\PageIndex{1}\)
In the above example, number of recombinant offspring was used to calculate map distance. However, the map distance can also be used to predict recombinant offspring. What percentage of fg / fg offspring will be produced from a cross between Fg / fG and fg / fg if loci F and G are 30 map units apart?
Solution
The homozygous recessive parent can only transmit an f g chromosome, so determine the percentage of f g gametes from the dihybrid parent. The parental chromosomes are F g and f G. The map distance (30 m.u.) is equal to the recombination frequency, so 30% of gametes will be recombinant, but there are two types of recombinants, so 15% will be F G and 15% will be f g. Therefore, we predict 15% of offspring will be f g / f g.
Exercise \(\PageIndex{2}\)
Two hypothetical SNPs in humans are 4 map units (m.u.) apart. The allele at SNP 1 can be A or T; the allele at SNP 2 can be C or G.
A male with genotype AG / TC and a female with genotype AC / AC have a child.

What is the probability they have a child with genotype AG / AC?
- Answer
-
The female will produce eggs with an AC chromosome. (Recombination still occurs in during meiosis in this female, but with or without recombination, the outcome is the same for these two SNPs.) To have chromosomes AG / AC, child must inherit the AG parental chromosome from the father.

The map distance (4 m.u.) indicates that 4% of gametes will be recombinant. In the male sperm, 4% of gametes will contain a recombinant (AC or TG) chromosome, and 96% of gametes will be parental: 48% of gametes will have the AG chromosome and 48% will have the TC chromosome. Therefore, the probability of an AG / A C child is 48%.
Query \(\PageIndex{1}\)
Query \(\PageIndex{2}\)
Query \(\PageIndex{3}\)
Double crossovers
Especially for large chromosomes, multiple crossover events can occur on the same chromosome. A double crossover occurs when fragments of the chromosome are exchanged in two places. The result of a double crossover is that the two ends of the chromosome are parental, but a region between the crossovers has been "swapped" for another sister chromatid sequence; this is depicted in the video.
Three factor crosses
Distances between multiple loci can be determined using three factor testcrosses. Again, we will cross a heterozygous parent to a parent homozygous recessive for all three genes. When solving three factor test crosses, remember that in the heterozygote the dominant and recessive alleles can be on the same or different chromosomes. You will know which chromosomes are parental because they will be the most abundant offspring from the testcross. In contract, the double crossover offspring will be the least abundant, because the double crossover events between the genes of interest are more rare than single crossovers.
Note: Steps for solving a three factor cross
- Identify the parental offspring (the most abundant).
- Identify the double crossover offspring (the least abundant).
- Determine which locus is in the middle (the one that "swaps places" in the double crossover).
- Determine the recombination frequency between one locus and the middle locus.
- Determine the recombination frequency between the other locus and the middle locus.
- Draw the genetic map.
Note that other factors that might influence recombination and double crossovers, such as the position along the chromosome or whether recombination at one site suppresses recombination nearby. These might make the number of observed recombinants different from expected, but we will not consider these factors at this time.
Exercise \(\PageIndex{3}\)
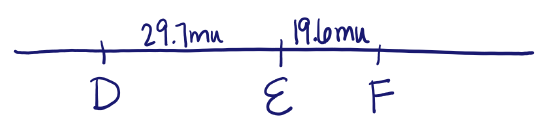
Consider three genes on the same chromosome, geneD, geneE and geneF. To determine the distance between and order of these three genes, you conduct a test cross between a heterozygote DEF / def and a def / def homozygous recessive. The offspring produced from the cross are shown in the table. What is the distance between the genes? Draw a map showing the position and distance between the three loci.
| Offspring class | Number of offspring |
| DEF | 84 |
| def | 86 |
| Def | 41 |
| dEF | 43 |
| DEf | 25 |
| deF | 27 |
| DeF | 5 |
| dEf | 5 |
- Answer
-
The distance between D and E is 29.7 m.u.; the distance between E and F is 19.6 m.u.