
2.4: Regeneration
- Page ID
- 27170

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Regeneration is the ability to replace lost or damaged body parts. This ability varies greatly among living things.
Plants
Plants can regenerate all body parts from precursor cells. Many trees, for example, can be cut off at the ground and, in due course, sprouts appear at the margins of the stump. These go on to develop new stems, leaves, and flowers. In the laboratory, entire plants can develop from a mass of undifferentiated cells growing in culture.

Figure \(\PageIndex{i}\): Carrot plant grown in a laboratory courtesy of Roy De Carava and Scientific American
For example, a fully-differentiated carrot root cell when grown in a suitable culture medium, begins to divide repeatedly, losing its differentiated structure as it does so. Then its descendants begin to differentiate, and they finally form all the organs of a mature carrot plant. The above photo shows a carrot plant that grew in a flask from fully-differentiated root cells that had been isolated and induced to undergo mitosis.
Invertebrate Animals
Sponges
Sponges can regenerate the entire organism from just a conglomeration of their cells.
Hydra
This cnidarian can also regenerate its entire body from cells. The cells that do the job are totipotent stem cells residing in the animal's body.
Planarians
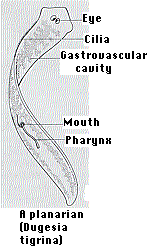
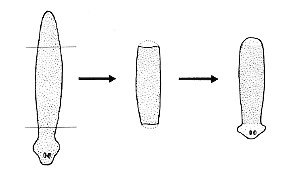
Figure \(\PageIndex{i}\): Dugesia and Dugesia regenaration
When some species of flatworms (left) are decapitated, they can regenerate a new head. Double-amputees can regenerate both a new head at the anterior surface and a new tail at the posterior surface (right). They do this by the proliferation and differentiation of the pluripotent stem cells (called neoblasts) that it retains in its body throughout its life.
How do the cells know whether to develop into a head or a tail? Thanks to the ease with which individual genes can be knocked out by RNA interference (RNAi), it has been shown that Wnt/β-catenin signaling dictates where the head and tail form.
- Blocking Wnt/β-catenin signaling by RNAi causes a head to form where a tail should (producing a two-headed animal) while
- blocking part of the β-catenin degradation complex (thus enhancing the pathway) causes a tail to develop where a head should (producing a two-tailed animal).
Thus it appears that the default pathway of neoblasts is to regenerate a head. In the amputated animal, a gradient of Wnt/β-catenin signaling extends from a high in the posterior — leading to the formation of a tail — and decreasing towards the anterior until the default pathway is no longer inhibited and a head can form.
Sea Stars (aka "Starfish')

Figure \(\PageIndex{i}\): Sea star regenerating an arm courtesy of Dr. Charles Walcott
These echinoderms can regenerate the entire organism from just one arm and the central disk. I have read that at one time oyster fishermen used to dredge up sea stars from their oyster beds, chop them up in the hope of killing them, and then dump the parts back overboard. They soon discovered to their sorrow the remarkable powers of regeneration of these animals.
Vertebrates
Newts and Salamanders

Figure \(\PageIndex{i}\): Salamander
These amphibians can regenerate a missing tail, legs, even eyes. This remarkable ability is particularly pronounced in the larval stage. For this reason, larval salamanders are favorites for doing research on regeneration. For example, cutting the tail off a larval salamander initiates the following sequence of events:
- A layer of epidermal cells grows over and covers the stump.
- A mass of undifferentiated cells — called the blastema — develops just beneath.
- Muscle and cartilage form in the regrowing tail.
- The notochord and spinal cord grow out into the regrowing tail.
- After a few weeks, a new, fully-functional and anatomically-correct tail is complete.
The Mechanism
For years, it has been unclear as to whether this regeneration depends on
- a population of pluripotent stem cells that have resided in the animal body prepared for such an event (as occurs in the hydra) or
- the dedifferentiation of specialized cells, e.g. muscle and cartilage cells, in the stump.
The answer appears to be both.
- Stem cells in the spinal cord migrate into the regrowing tail and differentiate into several cell types, including muscle and cartilage. Although the stem cells are ectoderm, they are able to develop into mesoderm.
- Muscle cells in the stump migrate into the blastema while
- reentering the cell cycle to produce thousands of descendants;
- dedifferentiate as they do so; that is, they lose the characteristic proteins, etc. of muscle cells.
- Even though there is as yet no sign of a tail, its final pattern is established during this process for if the blastema is removed and transplanted elsewhere, it will continue the process of regenerating a tail.
- Finally the cells of the blastema differentiate into all the cell types — nerve, muscle, cartilage, skin — used to build the regenerated tail.
Mammals
Don't we wish that we had the same powers of regeneration that salamanders do: able to regenerate a severed spinal cord or grow a new heart! But unfortunately, we cannot. We can regenerate some skin, a large amount of liver, and the very tips of fingers and toes. But that's about it. Just why we are so limited is not known (but is the subject of intense research). Much of the excitement surrounding research on stem cells is because of the hope that they may provide a means of regrowing damaged or lost tissues or even organs.
In contrast to the situation that appears to hold for salamanders, dedifferention of specialized cells does not appear to play a role in the formation of a blastema in mice. Instead, the various tissues — epidermis, hair follicles, sweat glands, neurons (all ectoderm) and muscle, bone, tendon, blood vessels (mesoderm) — that participate in regenerating the tip of an amputated mouse digit (finger or toe) develop from a diverse population of "adult" stem cells in the stump that retain their restricted developmental potential. You can read about the evidence for this in Rinkevich, Y., et al., Nature, 476, 409-413 (25 August 2011).
Genetic Control of Regeneration
A number of genes have been found to be implicated in regeneration. One of the most potent of these is Wnt.
- Injection of agents (e.g. antisense RNA molecules) that interfere with the Wnt/β-catenin pathway
- blocks limb regeneration in salamanders and, as we saw above,
- promotes head formation in regenerating planarians, while
- injection of agents that enhance the Wnt/β-catenin pathway
- enable chicks (that, like mammals, are normally incapable of regenerating limbs) to regenerate a wing;
- as well as enabling a regenerating planarian to form a tail where a head should go.
Contributors
-
John W. Kimball. This content is distributed under a Creative Commons Attribution 3.0 Unported (CC BY 3.0) license and made possible by funding from The Saylor Foundation.