
4.1: Meiosis
- Page ID
- 25729

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Learning Objectives
- Describe and draw the key events and stages of meiosis that lead to haploid gametes.
- Recall that homologous chromosomes separate during meiosis I (a reductional division) and that sister chromatids separate during meiosis II (an equational division).
- Compare mitosis and meiosis.
- Compare the processes of oogenesis and spermatogenesis in humans, including the chromosome complements of the gametes.
Most eukaryotes replicate sexually - a cell from one individual joins with a cell from another to create the next generation. For this to be successful, the cells that fuse must contain half the number of chromosomes as in the adult organism. Otherwise, the number of chromosomes would double with each generation! The reduction in chromosome number is achieved by the process of meiosis. In meiosis, there are usually two steps, Meiosis I and II. In Meiosis I homologous chromosomes segregate, while in Meiosis II sister chromatids segregate. Most multicellular organisms use meiosis to produce gametes, the cells that fuse to make offspring. Some single celled eukaryotes such as yeast also use meiosis.
Meiosis begins similarly to mitosis (a cell has replicated its chromosomes and grown large enough to divide), but requires two rounds of division. In the first, known as meiosis I, the homologous chromosomes separate and segregate. During meiosis II the sister chromatids separate and segregate. Note how meosis I and II are both divided into prophase, metaphase, anaphase, and telophase. After two rounds of cytokinesis, four cells will be produced, each with a single copy of each chromosome.
Meiosis is divided into two stages designated by the roman numerals I (one) and II (two). Meiosis I is called a reductional division, because it reduces the number of chromosomes inherited by each of the daughter cells. Meiosis I is further divided into Prophase I, Metaphase I, Anaphase I, and Telophase I, which are roughly similar to the corresponding stages of mitosis, except that in Prophase I and Metaphase I, homologous chromosomes pair with each other, or synapse, and are called bivalents. This is an important difference between mitosis and meiosis, because it affects the segregation of alleles, and also allows for recombination to occur through crossing-over, as described later. During Anaphase I, one member of each pair of homologous chromosomes migrates to each daughter cell (1N). Meiosis II resembles mitosis, with one sister chromatid from each chromosome separating to produce two daughter cells. Because Meiosis II, like mitosis, results in the segregation of sister chromatids, Meiosis II is called an equational division.
Meiosis I
In meiosis I replicated, homologous chromosomes pair up, or synapse, during the pachytene stage of prophase I, line up in the middle of the cell during metaphase I, and separate during anaphase I. For this to happen the homologous chromosomes need to be brought together while they condense during prophase I. These attachments are formed in two ways. Proteins bind to both homologous chromosomes along their entire length and form the synaptonemal complex (synapse means junction). These proteins hold the chromosomes in a transient structure called a bivalent. The proteins are released when the cell enters anaphase I, so that the homologous chromosomes can be separated.
Query \(\PageIndex{1}\)

Figure \(\PageIndex{1}\): Meiosis is a process in which a diploid cell divides into 4 haploid cells. At the end of Meiosis there are four genetically different cells. The diagram shows Meiosis as a non-cyclic process. (CC BY SA Ali Zifan via https://commons.wikimedia.org/wiki/File:Meiosis_Stages.svg)
Chromosome condensation during meiosis
As meiosis proceeds, chromatin becomes increasingly condensed. In some organisms, the DNA becomes so condensed that it appears as a spot of DNA instead of a line under the microscope. As you might expect from condensed chromatin, little transcriptional activity occurs during these stages of meiosis, so cells must produce the needed mRNAs in advance of meiosis.

Homologous Recombination
Within the synaptonemal complex during prophase 1, homologous recombination, or crossing over, occurs. These are places where DNA endonucleases break two non-sister chromatids in similar locations and then covalently reattach non-sister chromatids together to create a crossover between non-sister chromatids (4.1.1: Homologous recombination). This reorganization of chromatids will persist for the remainder of meiosis and result in recombination of alleles in the gametes.
Crossovers function to hold homologous chromosomes together during meiosis I so that they segregate successfully; they also cause the reshuffling of allele combinations to create genetic diversity, which can have an important effect on evolution.
Meiosis II
At the completion of meiosis I there are two haploid cells, each with one, replicated copy of each chromosome (1n). Because only one copy of each homolog is present, bivalents are not formed. In metaphase of meiosis II, the chromosomes will once again be brought to the middle of the cell, but this time it is the sister chromatids that will segregate during anaphase II.
After cytokinesis there will be four cells, each containing only one unreplicated chromosome of each type. Meiosis II resembles mitosis in that the number of chromosomes per cell is unchanged - both are equational cell divisions – but in meiosis II all four cells have different genetic composition. There will be allelic differences among the gametes.
Query \(\PageIndex{2}\)
Outcomes of meiosis
The outcome of meiosis is a cell or cells with half the number of chromosomes as the starting cell. But what combinations of chromosomes are possible?
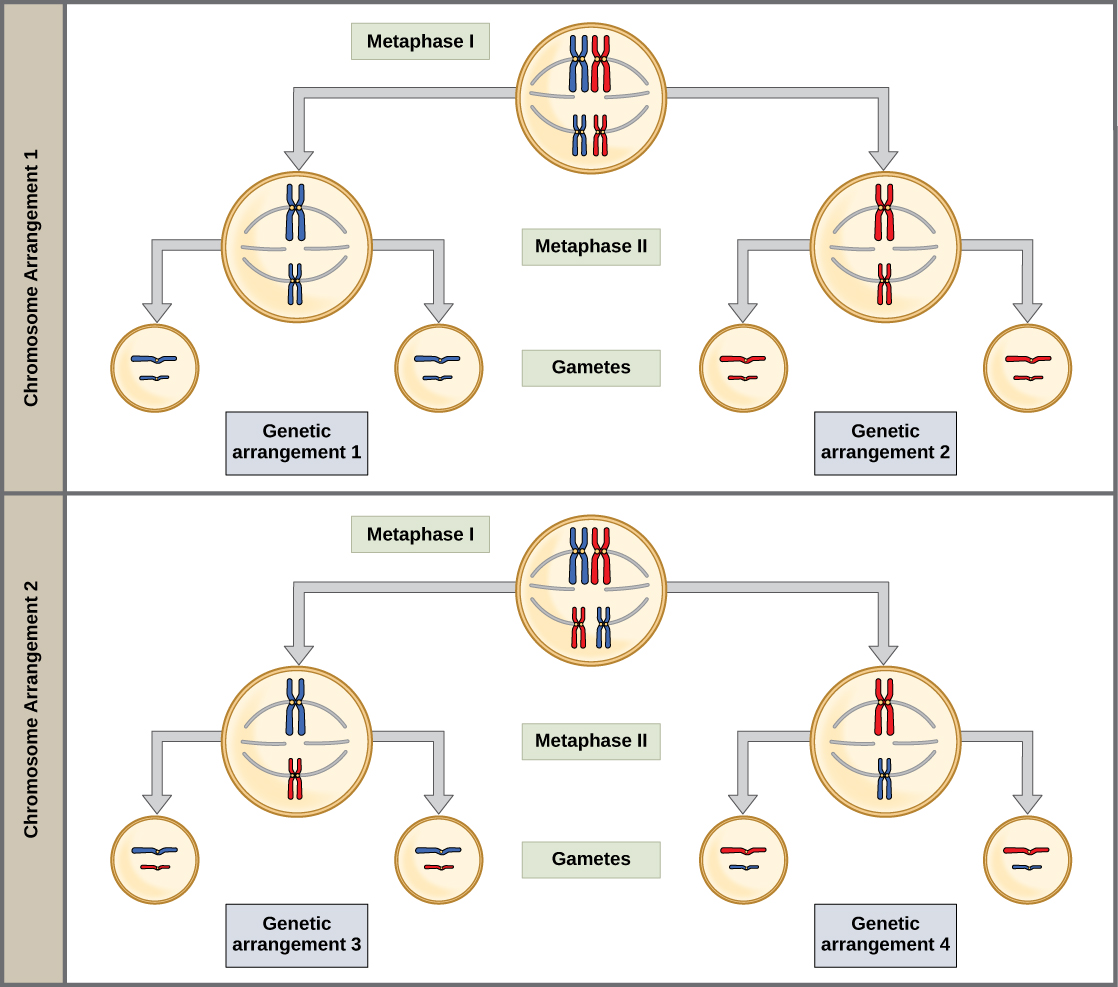
The homologous chromosomes separate during meiosis I, but the separation of the pairs of homologs is independent of other homologs. As an example in the figure below, for a cell with two pairs of chromosomes (2n=4), there can be four possible combinations of the four chromosomes. When we add recombination and additional pairs of chromosomes, there are almost infinite combinations of chromosomes in gametes.

Figure \(\PageIndex{2}\): Random, independent assortment during metaphase I can be demonstrated by considering a cell with a set of two chromosomes (n = 2). In this case, there are two possible arrangements at the equatorial plane in metaphase I. The total possible number of different gametes is 2n, where n equals the number of chromosomes in a set. In this example, there are four possible genetic combinations for the gametes. With n = 23 in human cells, there are over 8 million possible combinations of paternal and maternal chromosomes. (CC-BY OpenStax Figure_11_01_03.jpg)
{kind=link}
Watch the video below to follow how 4 pair of chromosomes are passed during meiosis I and II.
Video \(\PageIndex{1}\): How to count the number of chromosomes during the different phases of meiosis is explained. (www.youtube.com/watch?v=vCyiokyYkMw&t=3s)
Gamete maturation
In animals and plants, the cells produced at the end of meiosis need to mature before they become functional gametes.
Spermatogenesis
In most male animals, the four products of meiosis are called spermatids. They change morphology to develop tails and become functional sperm cells. In the meiotic steps of spermatogenesis, the cell divisions are equal, with the meiotic spindle aligned with the center of the cell, and the cells have equal amounts of cytoplasm, much like an average cell that has undergone mitosis. The streamlined, minimal-cytoplasm mature sperm is a product of post-meiotic differentiation, in which it gains the flagellar tail, and ejects most of its cytoplasmic material, keeping only some mitochondria to power the flagella and an acrosomal vesicle that contains the enzymes and other molecules needed to reach and fuse with (i.e. fertilize) a mature egg.

Oogenesis
In female animals, the gametes are oocytes. Each mature ovum (egg) will need to be as large possible to contain the maximum amount of cytoplasm including organelles, proteins, mRNAs, and nutrients to support the embryo after fertilization. To create large oocytes, only one of the four products of meiosis becomes an egg. The other three cells end up as tiny "disposable" cells called polar bodies, essentially little "packages" of extra DNA and very little cytoplasm. These cells are not viable and will eventually be degraded.
How can you make a really small cell?
The asymmetric distribution of cytoplasm in the first meiotic division for oocytes is due to the position of the meiotic spindle in the periphery of the cell rather than centered. During oogenesis, chromosomes do not line up in the middle of the cell during metaphase I or II. Because the center of the spindle determines the position of the contractile ring for cytokinesis, one small and one large cell are produced.

Timing of Spermatogenesis and Oogenesis
In addition to the differences in gamete size and number, in mammals the timing of meiosis differs between males and females. In males, germ cells are pre-meiotic at birth and do not enter meiosis until the onset of puberty. Mitosis maintains a population of precursor cells, so that sperm production can continue throughout adulthood. In females, germ cells enter meiosis I during embryonic development. These primary oocytes remain arrested ("stuck") in meiosis I until puberty. After this time, one primary oocyte per month (roughly for humans, depends upon cycle length for other mammals) completes meiosis I, enters meiosis II and is ovulated. Actually, meiosis II is only completed if the oocyte is successfully fertilized! The timing of meiotic arrest can differ between different species. For example, in the nematode C. elegans, oocytes are arrested in late prophase of meiosis I and only complete meiosis I and II rapidly after fertilization.
A note about plants
In plants, the products of meiosis reproduce a few times using mitosis as they develop into functional male or female gametes.
Fertilization
The purpose of gametes is to allow reproduction generation after generation. By uniting two gametes with half the number of chromosomes, the full chromosome number is restored each generation. Remember that homologous recombination and assortment of chromosomes create a genetically diverse population of gametes.
Fertilization restores the diploid number
oocyte (n) + sperm (n) = zygote (2n)
Errors in meiosis
Like any biological process, errors can occur during meiosis. If homologous chromosomes or sister chromatids are not correctly distributed during meiosis (known as nondisjunction), gametes can have too many or too few chromosomes (5.1: Changes in Chromosome Number).
Contributors and Attributions
Dr. Todd Nickle and Isabelle Barrette-Ng (Mount Royal University) The content on this page is licensed under CC SA 3.0 licensing guidelines.
- 15.8: Meiosis
- Stefanie Leacock, University of Arkansas-Little Rock