
7.5: Phloem Transport
- Page ID
- 74995

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Food and other organic substances (e.g., some plant hormones and even messenger RNAs) manufactured in the cells of the plant are transported in the phloem. Sugars (usually sucrose), amino acids and other organic molecules enter the sieve elements through plasmodesmata connecting them to adjacent companion cells. Once within the sieve elements, these molecules can be transported either up or down to any region of the plant moving at rates as high as 110 μm per second.
Two demonstrations:
- Girdling. Girdling is removing a band of bark from the circumference of the tree. Girdling removes the phloem, but not the xylem. If a tree is girdled in summer, it continues to live for a time. There is, however, no increase in the weight of the roots, and the bark just above the girdled region accumulates carbohydrates. Unless a special graft is made to bridge the gap, the tree eventually dies as its roots starve.

- The pictures below are autoradiographs showing that the products of photosynthesis are transported in the phloem.
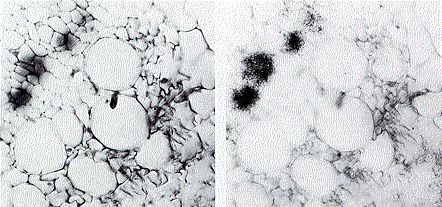
A cucumber leaf was supplied with radioactive water (3HOH) and allowed to carry on photosynthesis for 30 minutes. Then slices were cut from the petiole of the leaf and covered with a photographic emulsion. Radioactive products of photosynthesis darkened the emulsion where it was in contact with the phloem (upper left in both photos), but not where it was in contact with the xylem vessels (center). In the photomicrograph on the left, the microscope is focused on the tissue in order to show the cells clearly; on the right, the microscope has been focused on the photographic emulsion.
Some fruits, such as the pumpkin, receive over 0.5 gram of food each day through the phloem. Because the fluid is fairly dilute, this requires a substantial flow. In fact, the use of radioactive tracers shows that substances can travel through as much as 100 cm of phloem in an hour.
Mechanism that drives translocation of food through the phloem
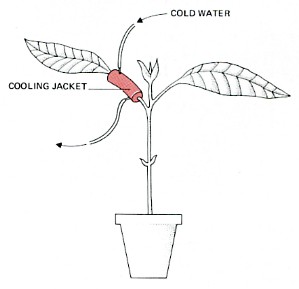
Translocation through the phloem is dependent on metabolic activity of the phloem cells (in contrast to transport in the xylem).
- Chilling its petiole slows the rate at which food is translocated out of the leaf (above).
- Oxygen lack also depresses it.
- Killing the phloem cells puts an end to it.
The Pressure-Flow Hypothesis
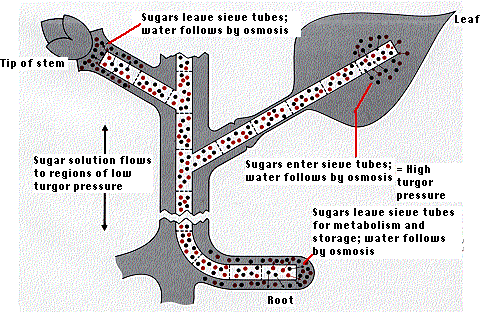
The best-supported theory to explain the movement of food through the phloem is called the pressure-flow hypothesis.
- It proposes that water containing food molecules flows under pressure through the phloem.
- The pressure is created by the difference in water concentration of the solution in the phloem and the relatively pure water in the nearby xylem ducts.
- At their "source" - the leaves - sugars are pumped by active transport into the companion cells and sieve elements of the phloem.
- As sugars (and other products of photosynthesis) accumulate in the phloem, water enters by osmosis.
In the figure, sugar molecules are represented in black, water molecules in red.)
- Turgor pressure builds up in the sieve elements (similar to the creation of root pressure).
- As the fluid is pushed down (and up) the phloem, sugars are removed by the cortex cells of both stem and root (the "sinks") and consumed or converted into starch.
- Starch is insoluble and exerts no osmotic effect.
- Therefore, the osmotic pressure of the contents of the phloem decreases.
- Finally, relatively pure water is left in the phloem, and this leaves by osmosis and/or is drawn back into nearby xylem vessels by the suction of transpiration-pull.
Thus it is the pressure gradient between "source" (leaves) and "sink" (shoot and roots) that drives the contents of the phloem up and down through the sieve elements.
Tests of the theory
1. The contents of the sieve elements must be under pressure.
This is difficult to measure because when a sieve element is punctured with a measuring probe, the holes in its end walls quickly plug up. However, aphids can insert their mouth parts without triggering this response.

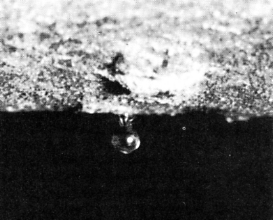
Left: when it punctures a sieve element, sap enters the insect's mouth parts under pressure and some soon emerges at the other end (as a drop of honeydew that serves as food for ants and bees).
Right: honeydew will continue to exude from the mouthparts after the aphid has been cut away from them.
2. The osmotic pressure of the fluid in the phloem of the leaves must be greater than that in the phloem of the food-receiving organs such as the roots and fruits. Most measurements have shown this to be true.
Transport of Messenger RNA (mRNA) through the Phloem
Plant scientists at the Davis campus of the University of California (reported in the 13 July 2001 issue of Science) have demonstrated that messenger RNAs can also be transported long distances in the phloem. They grafted normal tomato scions onto mutant tomato stocks and found that mRNAs synthesized in the stock were transported into the scions. These mRNAs converted the phenotype of the scion into that of the stock.