
Section 9.6: Complementation tests and Allelism
- Page ID
- 27249

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)– One, or more than one gene?
As explained earlier in this chapter, mutant screening is one of the beginning steps geneticists use to investigate biological processes. When geneticists obtain two independently derived mutants (either from natural populations or during a mutant screen) with similar phenotypes, an immediate question is whether or not the mutant phenotype is due to a loss of function in the same gene, or are they mutant in different genes that both affect the same phenotype (e.g., in the same pathway). That is, are they allelic mutations, or non-allelic mutations, respectively? This question can be resolved using complementation tests, which bring together, or combine, the two mutations under consideration into the same organism to assess the combined phenotype.
hypothetical example of purple flowers
The easiest way to understand a complementation test is by example (Fig.4.9). The pigment in a purple flower could depend on a biochemical pathway much like the biochemical pathways leading to the production of arginine in Neurospora (review in Chapter 1). A plant that lacks the function of gene A (genotype aa) would produce mutant, white flowers that looked just like the flowers of a plant that lacked the function of gene B (genotype bb). (The genetics of two loci are discussed more in the following chapters.) Both A and B are enzymes in the same pathway that leads from a colorless compound#1, thorough colorless compound#2, to the purple pigment. Blocks at either step will result in a mutant white, not wild type purple, flower.

Strains with mutations in gene A can be represented as the genotype aa, while strains with mutations in gene B can be represented as bb. Given that there are two genes here, A and B, then each of these mutant strains can be more completely represented as aaBB and AAbb . (LEARNING NOTE: Student often forget that genotypes usually only show mutant loci, however, one must remember all the other genes are assumed to be wild type.)
If these two strains are crossed together the resulting progeny will all be AaBb. They will have both a wild type, functional A gene and B gene and will thus have a pigmented, purple flower, a wild type phenotype. This is an example of complementation. Together, each strain provides what the other is lacking (AaBb). The mutations are in different genes and are thus called non-allelic mutations.
Now, if we are presented with a third pure-breeding, independently derived white-flower mutant strain, we won't initially know if it is mutant in gene A or gene B (or possibly some other gene altogether). We can use complementation testing to determine which gene is mutated. To perform a complementation test, two homozygous individuals with similar mutant phenotypes are crossed (Figure \(\PageIndex{10}\)).
If the F1 progeny all have the same mutant phenotype (Case 1 - Figure \(\PageIndex{10}\)A), then we infer that the same gene is mutated in each parent. These mutations would then be called allelic mutations - in the same gene locus. These mutations FAIL to COMPLEMENT one another (still mutant). These could be either the exact same mutant alleles, or different mutations in the same gene (allelic).
Conversely, if the F1 progeny all appear to be wild-type (Case 2 - Figure \(\PageIndex{10}\)B), then each of the parents most likely carries a mutation in a different gene. These mutations would then be called non-allelic mutations - in a different gene locus. These mutations do COMPLEMENT one another.
Note: For mutations to be used in complementation tests they are (1) usually true-breeding (homozygous at the mutant locus), and (2) must be recessive mutations. Dominant mutation CANNOT be used in complementation tests. Also, remember, some mutant strains may have more than one gene locus mutated and thus would fail to complement mutants from more than one other locus (or group).
| A. | 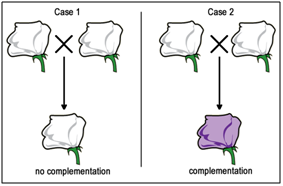 |
B. | 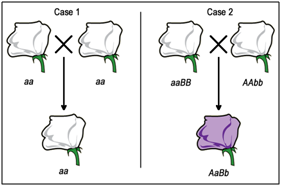 |
|
Figure \(\PageIndex{10}\)A – Observation: In a typical complementation test, the genotypes of two parents are unknown (although they must be pure breeding, homozygous mutants). If the F1 progeny all have a mutant phenotype (Case 1), there is no complementation. If the F1 progeny are all wild-type, the mutations have successfully complemented each other. |
Figure \(\PageIndex{10}\)B – Interpretation: The pure breeding, homozygous mutant parents had unknown genotypes before the complementation test, but it could be assumed that they had either mutations in the same genes (Case 1) or in different genes (Case 2). In Case 1, all of the progeny would have the mutant phenotype, because they would all have the same, homozygous genotype as the parents. In Case 2, each parent has a mutation in a different gene, therefore none of the F1 progeny would be homozygous mutant at either locus. Note that the genotype in Case 1 could be written as either aa or aaBB. (Original-Deyholos-CC:AN) |