
14.3C: DNA Replication in Eukaryotes
- Page ID
- 13293

DNA replication in eukaryotes occurs in three stages: initiation, elongation, and termination, which are aided by several enzymes.
- Describe how DNA is replicated in eukaryotes
Key Points
- During initiation, proteins bind to the origin of replication while helicase unwinds the DNA helix and two replication forks are formed at the origin of replication.
- During elongation, a primer sequence is added with complementary RNA nucleotides, which are then replaced by DNA nucleotides.
- During elongation the leading strand is made continuously, while the lagging strand is made in pieces called Okazaki fragments.
- During termination, primers are removed and replaced with new DNA nucleotides and the backbone is sealed by DNA ligase.
Key Terms
- origin of replication: a particular sequence in a genome at which replication is initiated
- leading strand: the template strand of the DNA double helix that is oriented so that the replication fork moves along it in the 3′ to 5′ direction
- lagging strand: the strand of the template DNA double helix that is oriented so that the replication fork moves along it in a 5′ to 3′ manner
Because eukaryotic genomes are quite complex, DNA replication is a very complicated process that involves several enzymes and other proteins. It occurs in three main stages: initiation, elongation, and termination.
Initiation
Eukaryotic DNA is bound to proteins known as histones to form structures called nucleosomes. During initiation, the DNA is made accessible to the proteins and enzymes involved in the replication process. There are specific chromosomal locations called origins of replication where replication begins. In some eukaryotes, like yeast, these locations are defined by having a specific sequence of basepairs to which the replication initiation proteins bind. In other eukaryotes, like humans, there does not appear to be a consensus sequence for their origins of replication. Instead, the replication initiation proteins might identify and bind to specific modifications to the nucleosomes in the origin region.
Certain proteins recognize and bind to the origin of replication and then allow the other proteins necessary for DNA replication to bind the same region. The first proteins to bind the DNA are said to “recruit” the other proteins. Two copies of an enzyme called helicase are among the proteins recruited to the origin. Each helicase unwinds and separates the DNA helix into single-stranded DNA. As the DNA opens up, Y-shaped structures called replication forks are formed. Because two helicases bind, two replication forks are formed at the origin of replication; these are extended in both directions as replication proceeds creating a replication bubble. There are multiple origins of replication on the eukaryotic chromosome which allow replication to occur simultaneously in hundreds to thousands of locations along each chromosome.
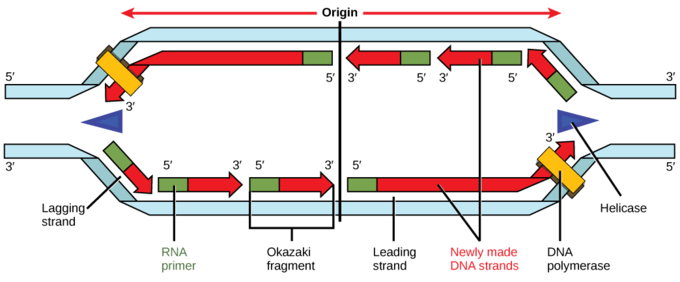
Elongation
During elongation, an enzyme called DNA polymerase adds DNA nucleotides to the 3′ end of the newly synthesized polynucleotide strand. The template strand specifies which of the four DNA nucleotides (A, T, C, or G) is added at each position along the new chain. Only the nucleotide complementary to the template nucleotide at that position is added to the new strand.
DNA polymerase contains a groove that allows it to bind to a single-stranded template DNA and travel one nucleotide at at time. For example, when DNA polymerase meets an adenosine nucleotide on the template strand, it adds a thymidine to the 3′ end of the newly synthesized strand, and then moves to the next nucleotide on the template strand. This process will continue until the DNA polymerase reaches the end of the template strand.
DNA polymerase cannot initiate new strand synthesis; it only adds new nucleotides at the 3′ end of an existing strand. All newly synthesized polynucleotide strands must be initiated by a specialized RNA polymerase called primase. Primase initiates polynucleotide synthesis and by creating a short RNA polynucleotide strand complementary to template DNA strand. This short stretch of RNA nucleotides is called the primer. Once RNA primer has been synthesized at the template DNA, primase exits, and DNA polymerase extends the new strand with nucleotides complementary to the template DNA.
Eventually, the RNA nucleotides in the primer are removed and replaced with DNA nucleotides. Once DNA replication is finished, the daughter molecules are made entirely of continuous DNA nucleotides, with no RNA portions.
The Leading and Lagging Strands
DNA polymerase can only synthesize new strands in the 5′ to 3′ direction. Therefore, the two newly-synthesized strands grow in opposite directions because the template strands at each replication fork are antiparallel. The “leading strand” is synthesized continuously toward the replication fork as helicase unwinds the template double-stranded DNA.
The “lagging strand” is synthesized in the direction away from the replication fork and away from the DNA helicase unwinds. This lagging strand is synthesized in pieces because the DNA polymerase can only synthesize in the 5′ to 3′ direction, and so it constantly encounters the previously-synthesized new strand. The pieces are called Okazaki fragments, and each fragment begins with its own RNA primer.
Termination
Eukaryotic chromosomes have multiple origins of replication, which initiate replication almost simultaneously. Each origin of replication forms a bubble of duplicated DNA on either side of the origin of replication. Eventually, the leading strand of one replication bubble reaches the lagging strand of another bubble, and the lagging strand will reach the 5′ end of the previous Okazaki fragment in the same bubble.
DNA polymerase halts when it reaches a section of DNA template that has already been replicated. However, DNA polymerase cannot catalyze the formation of a phosphodiester bond between the two segments of the new DNA strand, and it drops off. These unattached sections of the sugar-phosphate backbone in an otherwise full-replicated DNA strand are called nicks.
Once all the template nucleotides have been replicated, the replication process is not yet over. RNA primers need to be replaced with DNA, and nicks in the sugar-phosphate backbone need to be connected.
The group of cellular enzymes that remove RNA primers include the proteins FEN1 (flap endonulcease 1) and RNase H. The enzymes FEN1 and RNase H remove RNA primers at the start of each leading strand and at the start of each Okazaki fragment, leaving gaps of unreplicated template DNA. Once the primers are removed, a free-floating DNA polymerase lands at the 3′ end of the preceding DNA fragment and extends the DNA over the gap. However, this creates new nicks (unconnected sugar-phosphate backbone).
In the final stage of DNA replication, the enyzme ligase joins the sugar-phosphate backbones at each nick site. After ligase has connected all nicks, the new strand is one long continuous DNA strand, and the daughter DNA molecule is complete.
DNA Replication: This is a clip from a PBS production called “DNA: The Secret of Life.” It details the latest research (as of 2005) concerning the process of DNA replication.