
8.2: Case Study - The Evolution of Insect Wings
- Page ID
- 21547

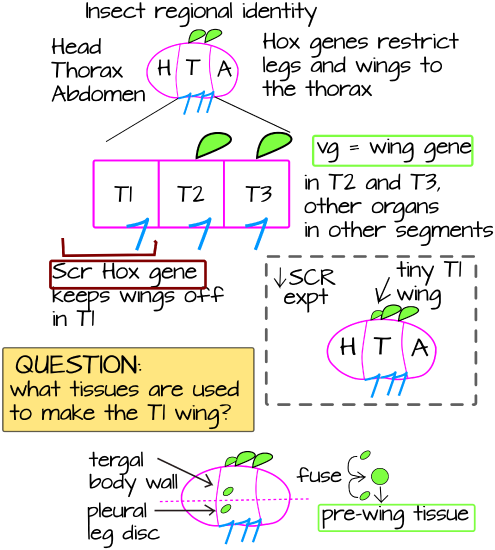
Insect wings are an incredibly important novelty associated with the radiation of the insects into one of the most diverse clades on the planet. They occupy land, water, and air and eat almost every food source imaginable. While their origin seems almost "out of the blue," careful developmental and paleontological studies have revealed key insights into their evolutionary history. Unlike insect legs, which form on the ventral side of the body, wings are dorsal appendages. This alone suggests a separate evolutionary origin from legs since they are not serially homologous. But, like legs, wings are found only on the thoracic segments - suggesting that Hox genes limit the expression of wing organizer genes. The insect body plan has three thoracic segments - T1, T2, and T3. Of these, only T2 and T3 build wings. In Drosophila, the T3 wings are reduced and form tiny halteres, which are proprioceptive organs. In beetles like Tribolium, which has been the focus of several wing development studies, the T2 wing forms an elytron (plural: elytra), a hard protective covering (Figure 1).

The Hox gene Scr represses wing development in T1, lowering Scr expresssion in Tribolium beetles gives an extra T1 outgrowth that resembles an elytron6. Looking more closely at the T1 tissue that forms the extra elytron in Scr- beetles, researchers in the Tomoyasu lab found that it is made up of two separate tissues, both of which normally express a gene called Vestigial6. Vestigial is normally expressed in early wing tissues (called "wing discs") but is also expressed in a few other tissues in the insect body. The Tomoyasu lab was very intrigued by the fact that two non-wing Vestigial expressing tissues were repressed from coming together to form a wing by Scr. These two tissues are called the pleural plate (the ventral-most tissue, referred to as "pleural") and the carinated margin (the dorsal-most tissue, referred to as "tergal"). The Tomoyasu lab hypothesized that wings evolved from these two body wall tissues fusing together and growing out of the body (Figure 2). Some evidence from this comes from comparing insects to crustaceans (Note here that I use "crustacean" to refer to all non-insect pancrustaceans but "crustacean" is not a true evolutionary clade).

Insects and crustaceans form a monophyletic clade called Pancrustacea and they share the same overall body plan. However, insects have derived limbs that are similar across the clade, while crustaceans in general have much more diversity. Ancestrally, pancrustaceans were marine organisms and had branched limbs, with one branch forming structures like gills. Insects obviously don't use gills, since they live on land and not in water, so the gill-forming structures in crustaceans are thought to form the pleural plates in insects. If the Tomoyasu hypothesis is correct, then insects have an ancestral gill forming structure joining up with a portion of the dorsal body wall (tergal location) to form a new appendage - the wing (Figure 2).
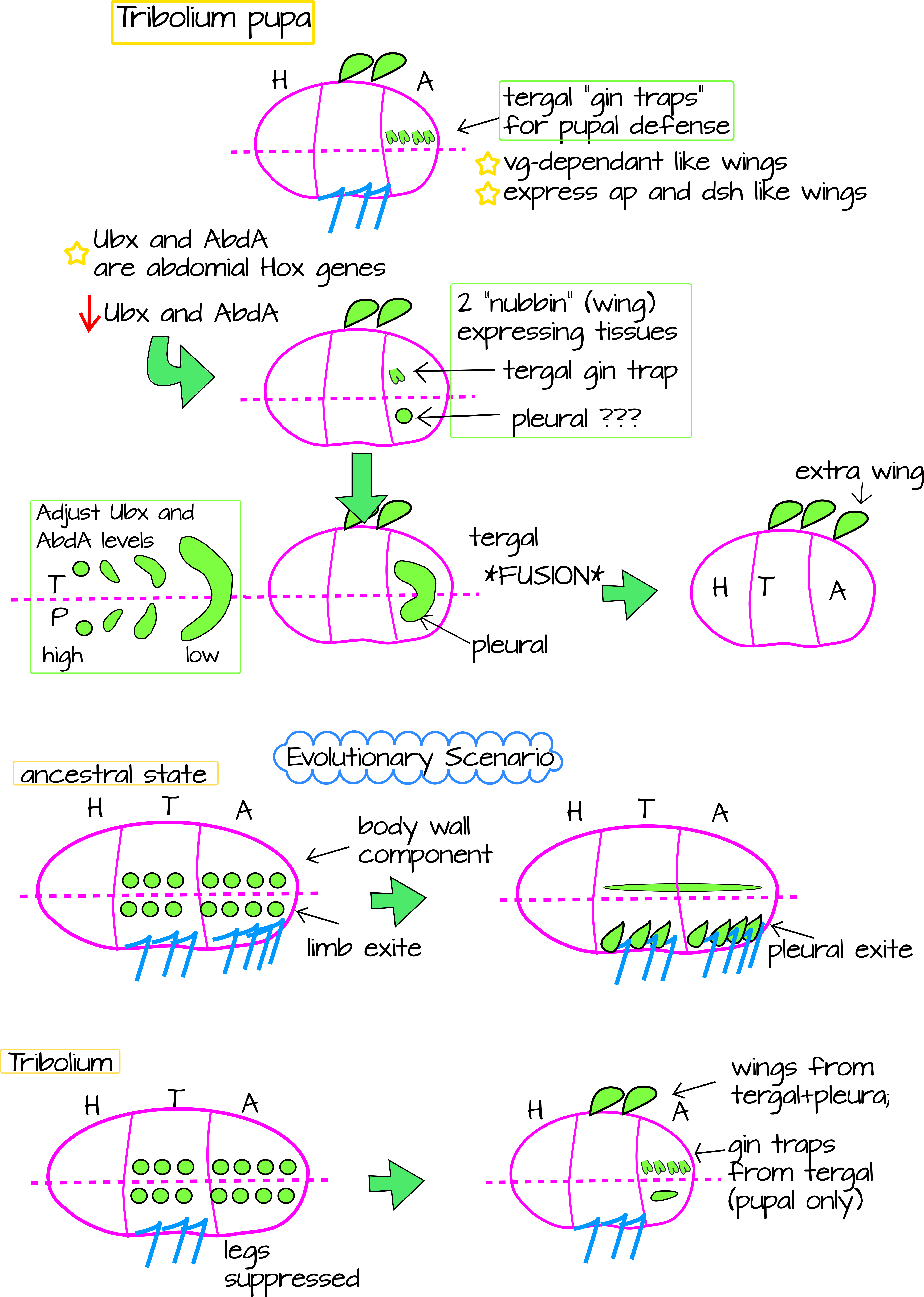
The Tomoyasu lab garnered more evidence for this when they looked for other serial homologs of wing tissue in abdominal segments. They found repeated Vestigial gene expression in dorsal body wall structures called "gin traps". These are defensive structures found in Tribolium pupae but disappear by adulthood. They express other wing-specifying genes like Vestigial, Apterous (Ap) Nubbin, and Disheveled (Dsh). Interestingly, they are formed on the abdomen, a part of the body in insects that is famous for not growing limbs. The lab also found a second tissue that expresses wing-specifying genes, the ventral carinated margin (Figures 1 and 2). The lab used the wing and gin-trap marker Nubbin to follow the development of both the carinated margin and the gin trap7.
When the researchers experimentally lowered levels of abdominal Hox genes Ubx and AbdA, they found that instead of gin traps, they got a fusion of the two Nubbin-expressing tissues. This fused tissue later formed a wing-like outgrowth in an abdominal segment (Figure 3). This suggest an evolutionary scenario wherein the pancrustacean ancestor had two Vestigal/Nubbin expressing tissues in its abdominal and thoracic segments. The tergal tissue would become part of the body wall and the pleural tissue would join the ventral legs to become the gill-branch. When insects moved to land, however, the gill-forming tissue was up for grabs since gills were no longer selected for. In the abdomen, the gill-forming tissue becomes part of the body wall, but in the thorax, it fuses with the Vestigial/Nubbin expressing tergal tissue to form wings. In the abdomen, where wing formation is suppressed, the tergal Vestigial/Nubbin tissue is free to take on a new fate. In the case of Tribolium, it forms a defensive structure in the abdomen, the pupal gin-traps (Figures 3 and Figure 4). In the thorax it forms the carinated margin. In this way, the wings are a novelty, but can also be thought of as serial homologs to the pleural plates+carinated margin in the thorax as well as to the gin-trap+pleural tissue in the abdomen.
Variations on a theme
Case study: bat wings
- http://genesdev.cshlp.org/content/22/2/141.full.
- http://journals.plos.org/plosgenetics/article?id=10.1371/journal.pgen.1005738#pgen.1005738.ref012.
- www.ncbi.nlm.nih.gov/pmc/articles/PMC1458926/.
Brand new stuff:
Case study: placenta