
8.2: Terrestrial Biomes
- Page ID
- 31619

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Because each biome is defined by climate, the same biome can occur in geographically distinct areas with similar climates (figure \(\PageIndex{a}\)). You can further explore the distribution of biomes using the HHMI Biome Viewer.
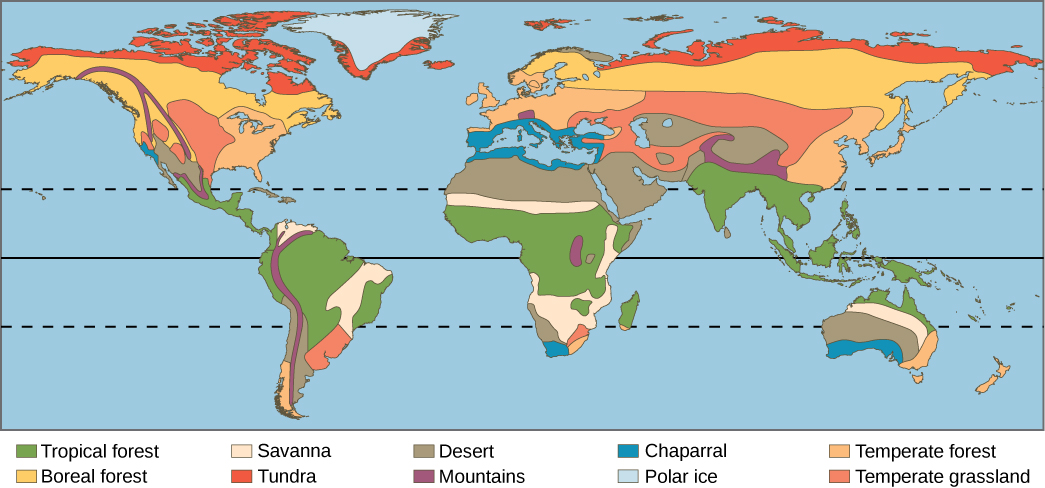
There are eight major terrestrial biomes: tropical rainforests, savannas, deserts, chaparral, temperate grasslands, temperate forests, taiga (boreal forests), and Arctic tundra.
Tropical Rainforests
Tropical rainforests, found in equatorial regions (figure \(\PageIndex{a}\)), are the most biodiverse terrestrial biome (figure \(\PageIndex{b}\)). In the Western Hemisphere, the tropical rain forest reaches its fullest development in the jungles of Central and South America. The trees are very tall and of a great variety of species. One rarely finds two trees of the same species growing close to one another. Most of the animals — mammals and reptiles, as well as birds and insects — live in the trees.

This biodiversity is under extraordinary threat primarily through logging and deforestation for agriculture. Tropical rainforests have also been described as nature’s pharmacy because of the potential for new drugs that is largely hidden in the chemicals produced by the huge diversity of plants, animals, and other organisms. The vegetation is characterized by plants with spreading roots and broad leaves. The broad-leafed trees are mostly evergreen, with leaves that persist throughout the year, but in tropical seasonal forests (see below), some are deciduous, with leaves falling off in the dry season.
The temperature and sunlight profiles of tropical rainforests are stable in comparison to other terrestrial biomes, with average temperatures ranging from 20oC to 34oC (68oF to 93oF). Month-to-month temperatures are relatively constant in tropical rainforests, in contrast to forests farther from the equator. This lack of temperature seasonality leads to year-round plant growth rather than just seasonal growth. In contrast to other ecosystems, a consistent daily amount of sunlight (11–12 hours per day year-round) provides more solar radiation and therefore more opportunity for net primary productivity (a measure of how carbon is stored by the photosynthesis that occurs in a community).
The annual rainfall in tropical rainforests ranges from 125 to 660 cm (50–200 in). Some tropical rainforests have relatively consistent rainfall whereas others (the tropical seasonal forests) have distinct wet months, in which there can be more than 30 cm (11–12 in) of precipitation, as well as dry months in which there are fewer than 10 cm (3.5 in) of rainfall. Nevertheless, the driest month of a tropical rainforest can still exceed the annual rainfall of some other biomes, such as deserts.
Tropical rainforests are characterized by vertical layering of vegetation and the formation of distinct habitats for animals within each layer. The vegetation is so dense that little light reaches the forest floor. On the forest floor is a sparse layer of plants and decaying plant matter. Above that is an understory of short, shrubby foliage. A layer of trees rises above this understory and is topped by a closed upper canopy—the uppermost overhead layer of branches and leaves. Some additional trees emerge through this closed upper canopy. These layers provide diverse and complex habitats for the variety of plants, animals, and other organisms. Many species of animals use the variety of plants and the complex structure of the tropical wet forests for food and shelter. Some organisms live several meters above ground, rarely descending to the forest floor.
Epiphytes
Epiphytes are plants that live perched on sturdier plants. They do not take nourishment from their host as parasitic plants do. Because their roots do not reach the ground, they depend on the air to bring them moisture and inorganic nutrients. Examples of epiphytes include many orchids, ferns (figure \(\PageIndex{c}\)), and bromeliads (members of the pineapple family like "Spanish moss").

Tropical Soils
While productivity is high in tropical rainforests, the soils themselves tend to be of very poor quality. Because of the high rainfall, nutrients are quickly washed out of the topsoil unless they are incorporated in the forest plants. As plant and animal debris falls to the ground, it is quickly decomposed because of the warmth and moisture there. Thus minerals are found mainly in the forest plants, not in the soil. When the plants are removed and cultivation attempted, the soils quickly lose fertility. The situation is made worse by the lack of humus. Additionally, the topsoil may be no thicker than 5 cm (~2 inches), and most of these soils have high iron and aluminum content. Once exposed to the sun, these soils quickly bake into a bricklike material that cannot be cultivated.
Savannas
Savannas have mostly perennial, herbaceous plants (non-woody plants that persist for more than one year) with scattered trees, and they are located in Africa, South America, and northern Australia. Savannas are usually hot, tropical areas with temperatures averaging from 24 °C to 29 °C (75 °F to 84 °F) and an annual rainfall of 10–40 cm (3.9–15.7 in). Savannas have an extensive dry season; for this reason, forest trees do not grow as well as they do in the tropical rainforest. As a result, grasses and forbs (herbaceous plants other than grasses) are dominant (figure \(\PageIndex{d-e}\)). Since fire is an important source of disturbance in this biome, plants have evolved well-developed root systems that allow them to quickly resprout after a fire. Depending on the region, large grazing animals like giraffes, zebras, and gazelles and their predators (cheetahs, lions, etc.) are found in savannas.


Deserts
Deserts exist between 15o and 30o north and south latitude. As discussed in the previous section (Climate and Biomes), deserts are frequently located on the downwind, or lee side, of mountain ranges due to the rain shadow effect. This is typical of the North American deserts, such as the Mohave and Sonoran deserts and the previously mentioned Great Basin Desert. Deserts in other regions (tradewind deserts), such as the Sahara and the Outback of Australia (both previously mentioned) as well as the Namib Desert in southwestern Africa and are dry because of the high-pressure, dry air descending at those latitudes.
Deserts are very dry; evaporation typically exceeds precipitation. Deserts are characterized by low annual precipitation of fewer than 30 cm (12 in) with little monthly variation and lack of predictability in rainfall. Some years may receive tiny amounts of rainfall, while others receive more. In some cases, the annual rainfall can be as low as 2 cm (0.8 in) in deserts located in central Australia (“the Outback”) and northern Africa.
Hot deserts (subtropical deserts) can have daytime soil surface temperatures above 60oC (140oF) and nighttime temperatures approaching 0oC (32oF). Cold deserts that experience freezing temperatures during the winter and any precipitation is in the form of snowfall. The largest of these deserts are the Gobi Desert in northern China and southern Mongolia, the Taklimakan Desert in western China, the Turkestan Desert, and the Great Basin Desert of the United States.
The low species diversity of this biome is closely related to its low and unpredictable precipitation. Despite the relatively low diversity, desert species exhibit fascinating adaptations to the harshness of their environment. Very dry deserts lack perennial vegetation that lives from one year to the next; instead, many plants are annuals that grow quickly and reproduce when rainfall does occur, then they die. Perennial plants in deserts are characterized by adaptations that conserve water: deep roots, reduced foliage, and water-storing stems (figure \(\PageIndex{f}\)). Seed plants in the desert produce seeds that can lie dormant for extended periods between rains. Most animal life in deserts has adapted to a nocturnal life, spending the hot daytime hours beneath the ground. The Namib Desert is the oldest on the planet, and has probably been dry for more than 55 million years. It supports a number of endemic species (species found only there) because of this great age. For example, the unusual gymnosperm Welwitschia mirabilisis the only extant species of an entire order of plants. There are also five species of reptiles considered endemic to the Namib.

Many of the animals in the desert (mammals, lizards and snakes, insects, and even some birds) are adapted for burrowing to escape the scorching heat of the desert sun. Many of them limit their forays for food to the night.
The net primary productivity of the desert is low. High productivity can sometimes be achieved with irrigation, but these gains are often only temporary because it leads to the buildup of salts and minerals. Large areas of formerly unproductive desert in the United States, Israel, and Egypt have been converted into fertile fields through irrigation. However, even the best irrigation water contains dissolved salts. Because the rainfall in deserts is so low, any water that does not immediately run off remains near the surface and is largely lost by evaporation. The salts it carries are left near the top of the soil. Their accumulation may make the soil so alkaline (basic) and so salty that it is prohibitive to agriculture (figure \(\PageIndex{g}\)). In the U. S., the situation is especially severe in the Great Basin because water flowing down from the mountains—bearing its load of dissolved salts—cannot flow on to the ocean but simply flows out onto the valley floors and evaporates.

Mediterranean Scrub (Chaparral)
The Mediterranean scrub, is also called chaparral or scrub forest and is found in California, along the Mediterranean Sea, and along the southern coast of Australia (figure \(\PageIndex{h}\)). The annual rainfall in this biome ranges from 65 cm to 75 cm (25.6–29.5 in) and the majority of the rain falls in the winter. Summers are very dry and many chaparral plants are dormant during the summertime.

The Mediterranean scrub vegetation is dominated by shrubs and is adapted to periodic fires, with some plants producing seeds that germinate only after a hot fire. The ashes left behind after a fire are rich in nutrients like nitrogen and fertilize the soil, promoting plant regrowth. Fire is a natural part of the maintenance of this biome. The trees in the chaparral are mostly oaks, both deciduous and evergreen. Scrub oaks and shrubs like manzanita and the California lilac (not a relative of the eastern lilac) form dense, evergreen thickets. All of these plants are adapted to drought by such mechanisms as waxy, waterproof coatings on their leaves. Many plants that thrive in one chaparral region are successfully cultivated in other chaparral regions. For example, vineyards, olives, and figs from their native Mediterranean and eucalyptus trees from Australia thrive in California. However, blue gum eucalyptus from Australia disrupts California ecosystems by changing the hydrology and outcompeting native plants.
Temperate Grasslands
Temperate grasslands are found throughout central North America, where they are also known as prairies (figure \(\PageIndex{i}\)), and in Eurasia, where they are known as steppes. Temperate grasslands have pronounced annual fluctuations in temperature with hot summers and cold winters. The annual temperature variation produces specific growing seasons for plants. Plant growth is possible when temperatures are warm enough to sustain plant growth, which occurs in the spring, summer, and fall.

Temperate grasslands have few trees except for those found growing along rivers or streams. The dominant vegetation tends to consist of grasses. The treeless condition is maintained by low precipitation, frequent fires, and grazing. Fires, which are a natural disturbance in temperate grasslands, can be ignited by lightning strikes. It also appears that the lightning-caused fire regime in North American grasslands was enhanced by intentional burning by humans. When fire is suppressed in temperate grasslands, the vegetation eventually converts to scrub and dense forests. Often, the restoration or management of temperate grasslands requires the use of controlled burns to suppress the growth of trees and maintain the grasses.
Annual precipitation ranges from 25.4 cm to 88.9 cm (10–35 in) and is concentrated in the summer and spring. In the plains of North America, the annual rainfall is sufficiently low (~50 cm, 20") that little or no rainfall percolates down to the water table. Calcium and other minerals are not carried below the reach of plant roots and so remain available for use. This keeps the pH and general fertility high. The grasses in undisturbed prairie are perennial. Their extensive root systems help prevent soil erosion, and the return of the season's above-ground growth to the topsoil returns minerals and provides humus to it. These advantages are diminished when annual grasses such as wheat and corn are grown instead and removed in the harvest.The self-restoring fertility of the soils of the plains states accounts for this region being the "breadbasket" of the nation (and other countries as well).
Temperate Forests
Temperate forests are the most common biome in eastern North America, Western Europe, Eastern Asia, Chile, and New Zealand (figure \(\PageIndex{i}\)). This biome is found throughout mid-latitude regions. Temperatures range between –30oC and 30oC (–22oF to 86oF) and drop to below freezing on an annual basis. These temperatures mean that temperate forests have defined growing seasons during the spring, summer, and early fall.

Temperate forests may be deciduous, mixed (with deciduous and evergreen trees), or coniferous (with evergreen conifers). Examples of temperate deciduous trees are beech, maple, oak, and hickory. The deciduous trees of temperate forests drop their leaves in the winter; thus, little photosynthesis occurs during the dormant winter period. (In other biomes, such as chaparral and tropical seasonal forests, trees or shrubs may drop their leaves during the summer or dry season.) Each spring, new leaves appear as temperature increases. Because of the dormant period, the net primary productivity of temperate forests is less than that of tropical rainforests. In addition, temperate forests show far less diversity of tree species than tropical rainforest biomes. Large stands dominated by a single species are common.
The deciduous trees of the temperate forests leaf out and shade much of the ground. However, more sunlight reaches the ground in this biome than in tropical rainforests because trees in temperate forests do not grow as tall as the trees in tropical rainforests. The soils of the temperate forests are rich in inorganic and organic nutrients compared to tropical rainforests. This is because of the thick layer of leaf litter on forest floors and reduced leaching of nutrients by rainfall. As this leaf litter decays, nutrients are returned to the soil. The leaf litter also protects soil from erosion, insulates the ground, and provides habitats for invertebrates and their predators. Deer, raccoons, and salamanders are characteristic inhabitants of temperate forests.
Precipitation is relatively constant throughout the year and ranges between 75 cm and 150 cm (29.5–59 in). Enough water falls on the soil so that much of it soaks deep into the ground and carries minerals with it. Such soils tend to be acidic and of low and (if unattended) diminishing fertility. Only by regular fertilization and liming (to restore calcium and raise pH) can productive agriculture be carried out in them. In the U.S., the soils east of the Appalachian Mountains tend to be of this sort.
Taiga
The taiga, also known as boreal forest in North America, is found roughly between 50° and 60° north latitude across most of Canada, Alaska, Russia, and northern Europe ((figure \(\PageIndex{i}\)). Boreal forests are also found above a certain elevation (and below high elevations where trees cannot grow) in mountain ranges throughout the Northern Hemisphere. This biome has cold, dry winters and short, cool, wet summers. The annual precipitation is from 40 cm to 100 cm (15.7–39 in) and usually takes the form of snow. Although precipitation is relatively low, the taiga is dotted with lakes, bogs, and marshes because the cold temperatures limit evaporation.

The long and cold winters in the boreal forest have led to the predominance of cold-tolerant cone-bearing plants. These are evergreen coniferous trees like pines, spruce, and fir, which retain their needle-shaped leaves year-round. Evergreen trees can photosynthesize earlier in the spring than deciduous trees because less energy from the sun is required to warm a needle-like leaf than a broad leaf. In the taiga, evergreen trees grow faster than deciduous trees. In addition, soils in boreal forest regions tend to be acidic with little available nitrogen. Leaves are a nitrogen-rich structure, and deciduous trees must produce a new set of these nitrogen-rich structures each year. Therefore, coniferous trees that retain nitrogen-rich needles in a nitrogen-limiting environment may have had a competitive advantage over the broad-leafed deciduous trees.
In North America, the moose is such a typical member that it has led to the name: "spruce-moose" biome. Deer, bears, and wolves are also found in the taiga. Before the long, snowy winter sets in, many of the mammals hibernate, and many of the birds migrate south.
The net primary productivity of taiga is lower than that of temperate forests and tropical rainforests. The aboveground biomass (mass of living organisms) of taiga is high because these slow-growing tree species are long-lived and accumulate standing biomass over time. Species diversity is less than that seen in temperate forests and tropical rainforests. Taiga lacks the layered forest structure seen in tropical rainforests or, to a lesser degree, temperate forests. The structure of the taiga is often only a layer of trees and the understory of low-lying plants and lichen. When conifer needles are dropped, they decompose more slowly than broad leaves; therefore, fewer nutrients are returned to the soil to fuel plant growth.
At the coldest parts of the taiga, the soil remains frozen year after year, forming a layer called permafrost. When the permafrost melts in unusually warm years, the trees are no longer supported by this hard surface and lean in different directions, forming a "drunken forest". At extreme latitudes, the trees of the taiga become stunted by the harshness of the subarctic climate. Where they finally disappear (above the "tree line"), the Arctic tundra begins.
Arctic Tundra
The Arctic tundra lies north of the subarctic boreal forests and is located throughout the Arctic regions of the Northern Hemisphere. Tundra also exists at elevations above the tree line on mountains. The average winter temperature is –34°C (–29.2°F), and the average summer temperature is 3°C–12°C (37°F –52°F). Plants in the Arctic tundra have a short growing season of approximately 50–60 days. However, during this time, there are almost 24 hours of daylight and plant growth is rapid. The annual precipitation of the Arctic tundra is low (15–25 cm or 6–10 in) with little annual variation in precipitation. And, as in the taiga, there is little evaporation because of the cold temperatures.
There is little species diversity, low net primary productivity, and low above-ground biomass. Like some of the taiga, the soils of the Arctic tundra may remain in a perennially frozen state, forming permafrost. The permafrost makes it impossible for roots to penetrate far into the soil and slows the decay of organic matter, which inhibits the release of nutrients from organic matter. The surface layer of soil above the permafrost melts in the brief summer, water is released and provides for a burst of productivity while temperatures and long days permit it. Because of these harsh growing conditions, the Arctic tundra is particularly sensitive to disturbances (disruptions that decrease biomass, such as heavy machinery passing through and squishing some of the plants).
Plants in the Arctic tundra are generally low to the ground and include low shrubs, sphagnum moss, grasses, lichens, and small flowering plants (figure \(\PageIndex{k}\)). Caribou feed on this growth as do vast numbers of insects. Swarms of migrating birds, especially waterfowl, invade the tundra in the summer to raise their young, feeding them on a large variety of aquatic invertebrates and vertebrates. As the brief arctic summer draws to a close, the birds fly south, and all but a few of the permanent residents, in one way or another, prepare themselves to spend the winter in a dormant state.

Attributions
Modified by Melissa Ha from the following sources:
- Terrestrial Biomes from Environmental Biology by Matthew R. Fisher (licensed under CC-BY)
- Biomes and Soil from Biology by John W. Kimball (licensed under CC-BY)
- Terrestrial Biomes from Biology 2e by OpenStax (CC-BY). Access for free at openstax.org.