
6.2: The Calvin Cycle
- Page ID
- 16122

The dark (carbon fixation) reactions vary depending on the type of plant. The most common set of carbon fixation reactions is found in C3-type plants, which are so named because the major stable intermediate is the 3-carbon molecule, glyceraldehyde-3-phosphate. These reactions, best known as the Calvin cycle (Figure \(\PageIndex{6}\)), fix CO2 onto the pentose, ribulose 1,5-bis-phosphate (RuBP).

The production part of the cycle begins with formation of RuBP from glyceraldehyde-3-phosphate. Then, the rate-limiting step occurs:
Ribulose 1,5-bisphosphate and CO2 are joined together by rubisco. Carboxylases are relatively slow enzymes as a family, and rubisco is one of the slowest. A 6-carbon intermediate is formed but it is unstable, and quickly breaks down to yield two molecules of 3-phosphoglycerate. Some familiar enzymes (from glycolysis, although this is happening in the stroma, not the cytoplasm) now come into play.
Phosphoglycerate kinase phosphorylates 3-PG to 1,3-bisphosphoglycerate. 1,3-BPG is then reduced by glyceraldehyde-3-phosphate dehydrogenase to form glyceraldehyde-3-P. This step requires the energy released from oxidation of NADPH. A small portion (1/6th) of the GAP that is made is then exported from the chloroplast and will be used to form more complex carbohydrates. However, the majority is recycled through the recovery phase of the Calvin cycle to regenerate NADP.
As if having a central enzyme that moves at a snail’s pace and needing to recycle the majority of its potential product was not bad enough, C3 plants also have to contend with the hijacking of rubisco for a competing, and energy-wasting, set of reactions known as photorespiration. Under conditions of low CO2 and high O2 in the local atmosphere, oxygen, instead of carbon dioxide, binds to rubisco and forms 3-PG and 2-phosphoglycolate from its reaction with RuBP. As detailed in Figure \(\PageIndex{7}\), the 2-phosphoglycolate is dephosphorylated to glycolate and transported out of the chloroplast. From there, it undergoes a series of reactions in the peroxisomes and mitochondria to transform it to 3-PG, which can then go in the chloroplast and participate in the Calvin cycle. Unfortunately for the cell, in the course of these reactions, NADH and ATP are used, thus lowering the energy availability inside the cell.
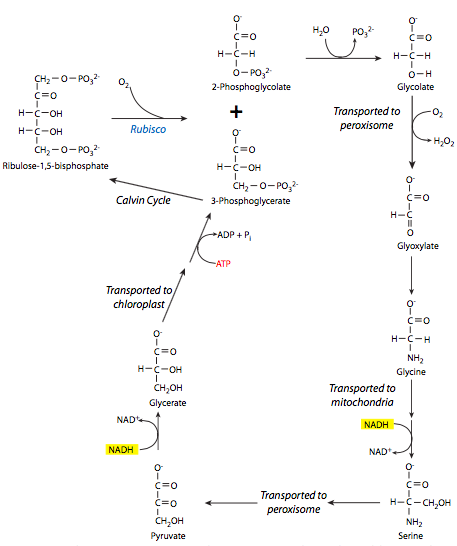
This is a particular problem in hot climates, because the oxygenase activity of rubisco increases more than the carboxylase activity as the temperature increases. This leads to an interesting side effect: in C3 plants, as the temperature rises and CO2 is outcompeted by O2 for rubisco binding, the stomata of the leaves need to remain open for longer in order to allow for acquisition of enough CO2 from the atmosphere. This in turn allows more water vapor from inside the cell to escape, leading to dehydration. C3 plants are thus at a competitive disadvantage in hot dry climates in comparison to plants that do not use rubisco for carbon fixation.
What about plants adapted to such climates? C4 plants, which include some grasses, corn, sugarcane, and weeds, utilize PEP carboxylase (which does not have the annoying photorespiratory capabilities of rubisco and a higher affinity for CO2) to fix carbon dioxide to PEP, making oxaloacetate. In an interesting twist, the oxaloactetate, after conversion to malate, is decarboxylated to yield CO2 again, which is fed to rubisco and the Calvin cycle.
PEP carboxylase actually fixes HCO3- to PEP rather than CO2 directly. The atmospheric CO2 is converted to the bicarbonate by carbonic anhydrase.
The C4 mechanism, also called the Hatch-Slack pathway, utilizes two sets of cells, an outer layer (mesophyll) that takes in air and fixes the CO2 to PEP and produces malate, and an inner layer of cells (bundle sheath) that takes the malate, and decarboxylates it for its rubisco enzyme. The two cells are connected via plasmodesmata (see Cell-cell Interactions chapter). Although energetically more expensive than carbon fixation by C3 plants in cooler climates, the C4 pathway overtakes C3 in efficiency as temperatures rise and photorespiration increases.
Variations of this pathway have been found in which aspartate is transported to the bundle-sheath cells instead of malate. After decarboxylation of the malate by malic enzyme (NAD-dependent in some species, NADP-dependent in others) to release the CO2 for rubisco, the resulting pyruvate is shuttled back to the mesophyll cell where it is phosphorylated by pyruvate-phosphate dikinase to generate PEP for re-entry into the C4 cycle.
Desert plants go one step further than C4 plants. Living in environments that are extremely hot and dry during the day, but relatively cool at night, many desert succulents (like cacti) are diurnal, and only open their stomata at night (when temperatures are significantly lower and water evaporates far more slowly) for CO2 gathering, which is then fixed via the CAM pathway to malate. Then in the daylight hours, CO2 is released from the malate and used in the Calvin cycle to generate carbohydrates.
The crassulacean acid metabolism (CAM) pathway is named for a carbon fixation pathway discovered in the Crassulaceae family of succulent plants including pineapples as well as various cactus species. It utilizes a similar biochemical mechanism as the C4 pathway, but occurs within a single photosynthetic cell. The major difference is that the CO2 is only taken in at night, and it quickly turned into malate, which is stored in vacuoles until daytime. The malate is then released and decarboxylated to provide the RuBP carboxylase (rubisco) with a steady stream of CO2 for fixation. Because there is such a rush of PEP carboxylase activity at night to fix the atmospheric CO2 to PEP, there is a high rate of starch breakdown to provide the glucose for glycolytic generation of PEP. Interestingly, as the malate is decarboxylated in the day, its product, pyruvate, can then be used to re-synthesize glucose (see gluconeogenesis section) and then starch.