
16.2: Viral Life Cycles
- Page ID
- 16192

Lytic “life” cycle of viruses
Viruses can interact with their hosts in two distinct ways: the lytic pathway and the lysogenic pathway. Some viruses are able to switch between the two pathways while others only use one. The distinguishing characteristic of the lytic life cycle is catastrophic death of the host cell by lysis and simultaneous release of viral particles. In Figure \(\PageIndex{2}\), the stages of the lytic pathway are depicted. In this case, a T4 bacteriophage (the term “phage” is used for bacterial viruses) is used as an example. In step 1, the virus attaches to the cell wall. In step 2, the virus injects its genetic material (dsDNA) into the cytoplasm of the bacteria. In step 3, the viral DNA is being replicated and the genes on the viral DNA are being transcribed and translated into viral proteins. Expression from the host genomic DNA is arrested. In step 4, viruses are assembled from the proteins and DNA. And finally, once the viral factory has used up the cell’s energy and material resources in making more viruses, it performs a final coup de grace, as cell is destroyed to free the viruses to exit and find more host cells. The T4 phage used in this example only undergoes this pathway and not the lysogenic pathway.
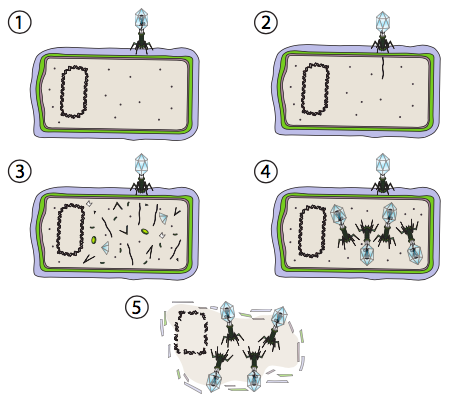
In eukaryotes, the mechanism is slightly more complicated by the nucleus. The DNA is transported into the nucleus, where the transcription and replication take place. Although the viral mRNA is transported out to the cytoplasm for translation as expected, the resulting capsid proteins are then imported back into the nucleus, where the virion particles are assembled. Lytic plant and animal viruses with RNA genomes can bypass the nucleus altogether, and the genome replication, protein synthesis, and particle assembly all occur in the cytoplasm.
The lytic pathway can produce a huge number of viral particles between infection and lysis, as many as several tens of thousands, for example from a rabies-infected cell. Therefore, this pathway is well suited for conditions in which potential host cells are plentiful. On the other hand, this is a waste of resources if there are relatively few potential hosts. Imagine a few bacteria that have floated off from the colony: if a phage infected a bacteria in the main colony, commandeering the bacteria to create thousands of viral particles, most of those particles would infect new hosts and make many thousands more soldiers in this viral army. But if the virus infected one of the breakaway bacteria, then once it killed its host by lysis, the viral particles would have few, if any, other potential hosts, and eventually all the viral particles just break down from various environmental conditions. What would a better survival strategy for the virus in such a situation?
The Lysogenic Pathway
A better option for some bacterial viruses is called the lysogenic pathway. The bacteriophage that have this option, as well as a lytic pathway, are known as temperate phage. In this pathway, the virus goes into dormancy by integrating into the host genome, and remaining transcriptionally quiescent until environmental conditions change and reflect a likelihood of more host cells to infect (Figure \(\PageIndex{4}\)).
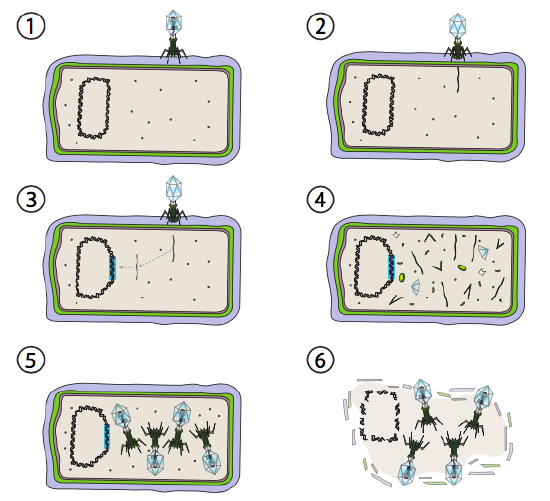
Lambda (λ) is an example of a temperate bacteriophage. The initial stages of infection and genome injection are the same as the lytic cycle, but under conditions that encourage lysogeny, the viral genome is integrated into the host genome in step 3. In λ integration into E. coli, this occurs by reciprocal recombination at a 15-base pair sequence known as the att λ site and is facilitated by the Int gene product. As long as the environmental conditions are not conducive to bacterial reproduction (and thus limited number of possible host cells), the viral genome remains mostly hidden and inactive. The only significant exception is a gene encoding a λ repressor that prevents the next step and keeps the virus dormant.
That next step is the excision of the λ phage DNA from the host chromosome, and subsequent replication and transcription of the viral DNA (Figure \(\PageIndex{4}\), Step 4). Then, like before, the final steps are assembly and accumulation of virions, and eventual breakdown of cellular structure and release of the viral particles.
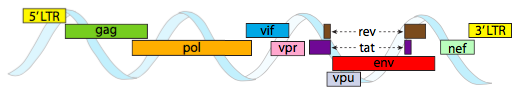
In order for packaging into the tight space constraints afforded by capsids, viral genomes must be highly economical. For example, the HIV genome (Figure \(\PageIndex{6}\)) has several genes that overlap.
Or, in the case of curtoviruses, ssDNA plant viruses (e.g. beet curly top virus), the genome not only has over- Rep lapping genes, it is even bi-directional (Figure \(\PageIndex{7}\)) encoding gene products in both strands of DNA after the ssDNA has been converted to dsDNA.
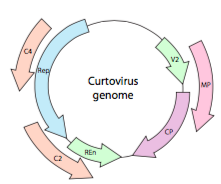
Given the need for economy, what genes are found in viruses? One of the most studied viral genomes, bacteriophage λ, contains genes encoding five transcriptional control proteins (which ones are expressed depends on whether the phage is in a lysogenic or lytic mode), a binding protein that controls degradation of a transcriptional activator, 17 capsid proteins, an excisionase that controls excision and insertion of the phage genome in the host genome, an integration protein that inserts the phage genome into the host’s, and 3 genes participating in lysis of the host cell.
The HIV genome depicted above is much smaller than λ, at around 9 kilobases compared to 48 kb, but again, the theme is to use cellular proteins when possible, and encode viral genes if necessary. So, gag encodes capsid proteins, pol encodes reverse transcriptase, integrase, and HIV protease (which cleaves the gag and pol gene products into their functional proteins), vif acts against a common host cell antiviral enzyme, vpr regulates nuclear import, tat strongly increases transcription of HIV genes, rev exports viral RNA from the nucleus, vpu is needed for budding of particles from the host, env encodes viral envelope glycoproteins, and nef promotes survival of infected cells. The LTR regions are very strong promoters to drive high expression of these genes.
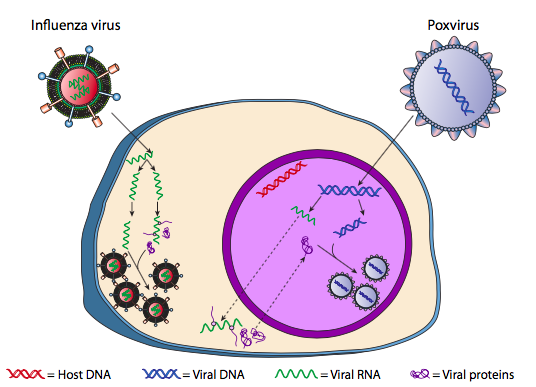
Although it is not referred to as lysogeny, some animal viruses can behave similarly. The most prominent example is the Baltimore Class VI viruses - commonly known as retroviruses, one of which is HIV. The path of a retrovirus through a eukaryotic host cell is depicted below ( g. 5).
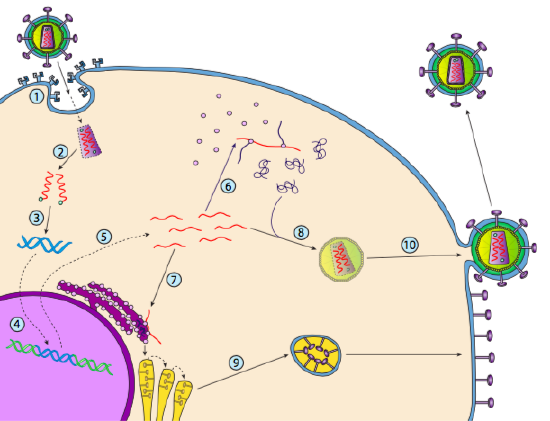
HIV has an envelope, which is studded with transmembrane proteins that are recognized by the host cell, binding the virus to the cell surface and initiating receptor-mediated endocytosis (1). After the endocytosis, the membrane envelope of the virion and the vesicular membrane fuse to release the capsid and its contents (2). After the capsid dissociates in the cytoplasm, the two strands of viral RNA are released along with a special polymerase: reverse transcriptase, which reads an RNA template and synthesizes DNA. Reverse transcriptase also uses that new DNA to synthesize a complementary DNA strand so that it eventually produces a double- stranded DNA version of the viral genome (3). This viral dsDNA is transported into the nucleus where it integrates into the host genome using another viral protein, integrase
(4). The integrated viral DNA is called a provirus. The provirus can lay dormant, but if it is activated, then it is transcribed and the resulting viral RNA is transported out of the nucleus (5). Some of the viral RNA encodes enzymes like reverse transcriptase and integrase, or capsid proteins, all of which are made in the cytoplasm (6), but some encode membrane bound glycoproteins, which are translocated into the ER (7) and eventually processed through the Golgi and incorporated into the plasma membrane (9). Once the virion has been assembled (8), it binds to the viral transmembrane proteins, nucleating an exocytic “vesicle” (10) which is the virion complete with viral envelope.
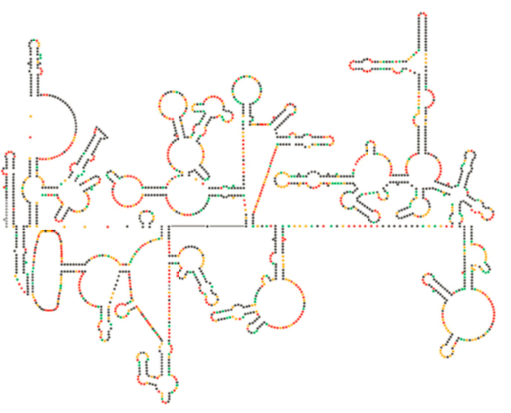
Recent structural examination of the HIV genome suggests that the structure of HIV RNA itself may play a significant role in its propagation inside of host cells. Figure \(\PageIndex{8}\), from Watts et al, Nature 460:711-716, 2009, shows a predicted secondary structure of the genome. The authors suggest that the RNA structure actually may interact with ribosomal elongation to control the folding of the viral proteins. They also postulate the extension of this argument to include important genetic information encoded not just in the nucleotide sequence, but the secondary structure and tertiary structure of any RNA virus.
In considering viruses with respect to the rest of this integrative chapter, there are two overriding ideas to keep in mind. First, viral survival is based on numbers: it needs to make huge numbers of its components to cast as wide a net for new host cells as possible. To do this, viral promoters are usually much stronger than host cell promoters, simultaneously driving more viral gene expression while preventing host gene expression (by dedicating cellular resources to virus production). Second, because of fast generation times, the rate of viral mutation and evolution is far faster than normal eukaryotic genomes. In addition, if the virus uses its own polymerase (such as reverse transcriptase or RNA replicase), the mutation rate rises even more because there is no error-checking by viral polymerases.