
7.4: Photosynthesis
- Page ID
- 16453

Chemically, photosynthesis is the reverse reaction of respiration. Compare the two reactions:
\[C6H12O6 + 6O2 ⇔ 6CO2 + 6H2O\] (DGo = -687Kcal/mole)
\[6CO2 + 6H2O ⇔ C6H12O6 + 6O2\] (DGo = +687Kcal/mole)
If respiration (reaction 1) is the complete oxidation of glucose to H2O and CO2, then photosynthesis (reaction 2) is the reduction of CO2 using electrons from H2O. Photosynthesis is thus an endergonic reaction. During photosynthesis, sunlight (specifically visible light), fuels the reduction of CO2 (summarized below).

Photosynthesis began in the absence of oxygen; it came before oxygenic respiration on earth. Increasing oxygen in the atmosphere led to selection of oxygenic respiratory pathways (the Krebs cycle, electron transport and oxidative phosphorylation). When we look at photosynthesis in some detail, we will see that photosynthesis and respiration have electron transport-ATP synthesizing systems with similar features. This suggests that they share a common evolutionary ancestry. Elsewhere, we will consider what a common ancestral system might have looked like. Two biochemical pathways make up photosynthesis:
· Light-dependent reactions that use visible light energy to remove electrons from water, reduce electron carriers, pump protons and make ATP;
· Light-independent reactions that use ATP to transfer electrons from the reduced electron carriers to CO2 to synthesize glucose.
The two pathways are summarized below.

A. The Light Dependent Reactions
Colored substances contain pigments that reflect the colors that we see and at the same time, absorb all the other colors of visible light. Early studies asked which plant pigments absorbed the light that allowed (we say supported) photosynthesis. Chlorophyll, the abundant pigment we see in plant tissues is actually two separate green pigments, chlorophyll a and chlorophyll b. One might therefore predict that light absorbed by chlorophyll will support photosynthesis, but light absorbed by other pigments in plant cells would not.
The experiment to test this hypothesis is illustrated below.

The action spectrum of photosynthesis below plots the results of this experiment.

The spectrum shows that all wavelengths of visible light energy support photosynthesis. In addition, other experiments revealed that radiation other than visible light (e.g., ultraviolet and infrared light) do not support photosynthesis. One can conclude that chlorophylls alone are likely not the only pigments to support photosynthesis.
Chlorophylls are easily purified from leaves. The graph below shows an average absorbance spectrum for of chlorophylls. The absorbance of chlorophyll a and chlorophyll b are slightly different, but center at wavelengths at 450 nm and 675 nm.
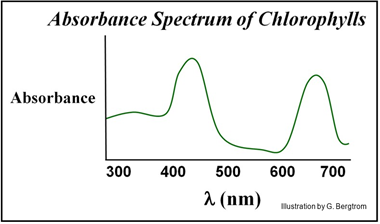
We can conclude from the absorbance spectra that chlorophylls do support photosynthesis, but that, indeed, they are not alone in doing so. Chlorophylls alone do not account for the action spectrum of photosynthesis! Clearly, other pigments absorbing elsewhere in the visible spectrum also support photosynthesis. Of course, we knew that leaves and other photosynthetic plant tissues contained a variety of different pigments, many of which we see as fall colors. All of these pigments (including chlorophylls) are found in the chloroplasts, the organelles that conduct photosynthesis in plants. Examine the structure of chloroplasts in the electron micrographs below.

The visible light absorbance spectra of three different kinds of plant pigments shown below do coincide with the action spectrum of photosynthesis. This implies that absorption of light by those pigments is responsible for photosynthesis.

Carotenoids, chlorophyll b and other accessory pigments participate in capturing light energy for photosynthesis. Two clusters of pigments capture light energy. These reaction centers are part of photosystems 1 and photosystem 2 on thylakoid membranes of chloroplasts. Johann Deisenhofer, Robert Huber and Hartmut Michel first determined the 3D structure of a bacterial reaction center. Then they and unraveled the relationship between the structure of the proteins in the center and the membrane in which they were embedded. For this, they shared the 1988 Nobel Prize in Chemistry.
The activities of Photosystem I are animated at Photosystem 1 Action. You should see light (a photon) excite electron (e-) pairs excited from Photosystem I pigments that then transfer their energy from pigment to pigment, ultimately to chlorophyll a P700. The impact of the electron pair then excites a pair of electrons from chlorophyll a P700. This e- pair is captured by a photosystem I (PSI) e- acceptor. Next, the reduced PSI acceptor is oxidized as electrons move down a short ETC, eventually reducing NADP+ to NADPH. Electrons on NADPH will eventually be used to reduce
CO2 to a carbohydrate. So far, so good! But that leaves an electron deficit in Photosystem I. The Z-Scheme illustrated below follows electrons taken from water (absorbed through roots) into photosystem II (PSII), which will replace those missing from PSI.

Let’s summarize the flow of electrons from water through the Z-scheme. Light excites an e- pair from the P680 form of chlorophyll a in PSII. A PSII electron acceptor in the thylakoid membrane, identified as pheophytin, captures these electrons. An electron transport chain oxidizes the pheophytin, transferring e- pairs down to PSI. Some of the free energy released pumps protons from the stroma into the space surrounded by the thylakoid membranes. The gradient free energy fuels ATP synthesis as protons flow back into the stroma through a chloroplast ATP synthase. The link at Action in the Z-Scheme animates the entire Z-Scheme, showing first how PSI electrons reduce NADP+ and then how PSII electrons replace missing PSI electrons, making ATP along the way. The oxygen released by splitting water ends up in the atmosphere.
B. Cyclic Photophosphorylation
The Z-Scheme does not in fact make enough ATP to power the Calvin Cycle. But when the need for ATP exceeds the capacity of the tissues to make sugar, the photosynthetic apparatus can take a time-out, resorting to Cyclic Photo- phosphorylation for a while. Cyclic Photophosphorylation simply takes electrons excited to the PSI electron acceptor, and instead of sending them to NADP+, deposits them on PC (plastocyanin) in the electron transport chain between PSII and PSI. These electrons then flow down this ‘long line’ of the Z, right back to PSI, releasing their free energy to make ATP. In light, the electrons just go up and around, hence the name Cyclic Photophosphorylation. The path of electrons is shown below and animated at Action in Cyclic Photophosphorylation.

C. The Light-Independent (“Dark”) Reactions
1. The Dark Reactions of C3 Photosynthesis
As we have seen, the light-dependent reactions of photosynthesis require light energy and water and generate O2, ATP and NADPH. In the light-independent (or ‘dark’) reactions, the ATP and NADPH will provide free energy and electrons (respectively) for carbon fixation (the reduction of CO2 to make carbohydrates). CO2 enters photosynthetic tissues through stomata. Stomata are pores in leaves that can be open or closed, depending on light, temperature conditions and water availability. In addition to allowing CO2 into photosynthetic tissues, stomata also function in transpiration, which allows excess water in cells to leave the plants by transpiration (sometimes called evapotranspiration). C3 photosynthesis is the mechanism of carbon fixation in most plants, so called because its first carbohydrate product is a 3-C molecule, 3-phosphoglyceric acid (3-PG). You should recognize 3-PG; it is also a glycolytic intermediate. The Calvin Cycle is the most common dark reaction pathway. For its discoverer, M. Calvin received the Nobel Prize in Chemistry in 1961. The Calvin Cycle is shown below.

Check the animation at Action in the Calvin Cycle.
Each carbon dioxide entering the Calvin cycle is "fixed" to a 5-carbon ribulose bisphosphate molecule (RuBP), catalyzed by the enzyme RuBP carboxylase- oxygenase, or Rubisco for short. The expected 6-C molecule must be quickly split into two 3-C molecules since it has not been detected as an intermediate to date! The first detectable products are two molecules of 3-PG. Each 3-PG is in turn reduced to glyceraldehyde-3-phosphate (G-3-P, another familiar molecule).
The cycle regenerates the RuBP and produces glucose. Perhaps the easiest way to see this is to imagine the cycle going around 12 times, fixing 12 molecules of carbon dioxide, as shown in the link above. Two of the G-3-P molecules are linked together to make a single 6-C molecule of glucose (which is polymerized into starch for storage in daylight). That leaves 10 molecules of G-3-P, for a total of 30 carbons. The latter part of the cycle will regenerate 5 molecules of new RuBP, accounting for our 30 carbons!
2. Photorespiration
There are times that even plants in temperate environments suffer prolonged hot, dry spells. Perhaps you have seen a lawn grow more slowly and turn brown after a dry heat wave in summer, only to grow and re-green after the rains resume. C3 plants resort to photorespiration during drought and dry weather, closing their stomata to conserve water. Under these conditions, CO2 can’t get into the leaves… and O2 can’t get out! As CO2 levels drop and O2 rise in photosynthetic cells, the Calvin Cycle slows down. Instead of fixing CO2, the enzyme Rubisco now catalyzes “O2 fixation” using its oxygenase activity. The combination of RuBP with O2 splits RuBP into a 3-carbon and a 2-carbon molecule: 3-phosphoglyceric acid (3-PG) and phosphoglycolate respectively. The reaction is shown below.

Not only does photorespiration result in only one 3-carbon carbohydrate (compared to two in the Calvin Cycle), but the phosphoglycolate produced is cytotoxic (not healthy for cells!). Removing the phosphate and metabolizing the remaining glycolic acid costs energy. Therefore, photorespiration can only be sustained for a short time. On the other hand, plants that have adapted to live in hot arid environments all the time have evolved one of two alternate pathways. One is the CAM (Crassulacean Acid Metabolism); the other is the C4 pathway. Each is an alternative to C3 carbon fixation.
3. The CAM Photosynthetic Pathway
Crassulacean acid metabolism (CAM) was discovered in the Crassulaceae. These are succulents like sedum (a common ground cover), cactuses and jade plants, and some orchids. The CAM pathway was selected in evolution to allow plants to conserve water, especially during the high daytime temperatures. Stomata in chlorenchymal (mesophyll) leaf cells close during the day to minimize water loss by transpiration. The stomata open at night, allowing plant tissues to take up CO2. CAM plants fix CO2 by combining it with PEP (phosphoenol pyruvate). This eventually produces malic acid that is stored in plant cell vacuoles. By day, stored malic acid retrieved from the vacuoles splits into pyruvate and CO2. The CO2 then enters chloroplasts and joins the Calvin Cycle to make glucose and the starches. The CAM pathway is shown below.

In sum, CAM plant mesophyll cells
1. open stomata to collect, fix and store CO2 as an organic acid at night.
2. close stomata to conserve water in the daytime.
3. re-fix the stored CO2 as carbohydrate using the NADPH and ATP from the light reaction the next the day.
4. The C4 Photosynthetic Pathway
C4 refers to malic acid, the 4-carbon end product of CO2 fixation. In this regard, the C4 pathway is the same as in CAM metabolism! In both pathways, PEP carboxylase is the catalyst of carbon fixation, converting phosphoenol pyruvate (PEP) to oxaloacetate (OAA). The OAA is then reduced to malic acid, as shown below.
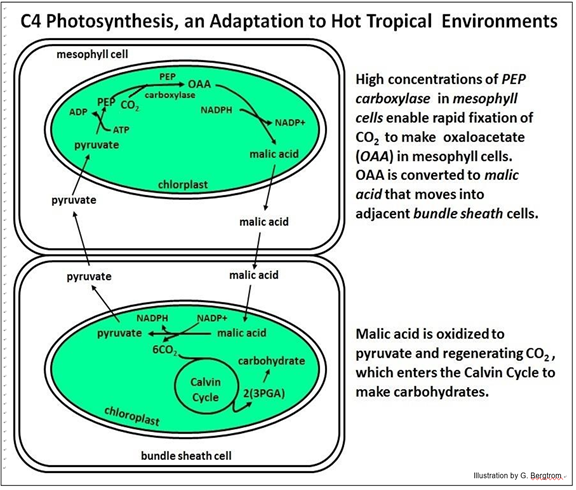
C4 metabolism diverges from CAM pathway after malic acid formation. PEP carboxylase catalysis is rapid in C4 plants, in part because malic acid does not accumulate in the mesophyll cells. Instead, it is rapidly transferred from mesophyll to adjacent bundle sheath cells, where it enters chloroplasts. The result is that C4 plants can keep stomata open for CO2 capture (unlike CAM plants), but closed at least part of the day to conserve water. The 4-carbon malic acid is oxidized to pyruvate (three carbons) in the bundle sheath cell chloroplasts. The CO2 released enters the Calvin cycle to be rapidly fixed by Rubisco. Of course, this system allows more efficient water use and faster carbon fixation under high heat, dry conditions than does C3 photosynthesis. Corn is perhaps the best-known C4 plant!
By the way, can you recognize several more intermediates common to respiration and the light-independent photosynthetic reactions?