
16.4: How Membrane Proteins are Held in Membranes
- Page ID
- 16516

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
The hydrophobic domain of integral membrane proteins consists of one or more alphahelical regions that interact with the hydrophobic interior of the membranes. Hydrophilic domains tend to have more tertiary structure with hydrophilic surfaces, and so face the aqueous cytosol and cell exterior. Two trans-membrane proteins are cartooned below.
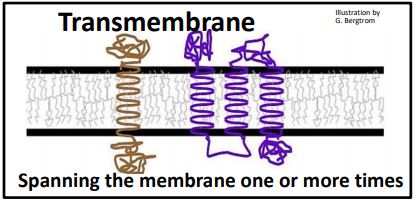
The protein on the left crosses the membrane once, while the one on the right crosses the membrane three times. How a transmembrane protein inserts into the membrane during synthesis dictates the locations of its N- and C-terminus. Transmembrane proteins can in fact cross a membrane more than once, which also determines the location of its N- and C-termini. N-terminal end of a plasma membrane polypeptide always ends up exposed to the outside of the cell. The alpha helical domains that anchor proteins in membranes are mostly non-polar and hydrophobic themselves. As an example, consider the amino acids in the alpha-helical domain of the red blood cell protein glycophorin A, a membrane protein that prevents red blood cells from aggregating, or clumping in the circulation. One glycophorin A polypeptide with its hydrophobic trans-membrane alpha helix is cartooned below. Glycophorin A monomers pair to form dimers in the plasma membrane.
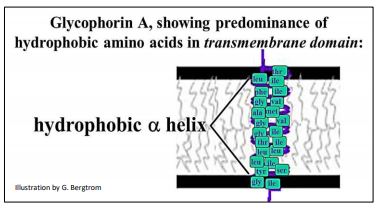
Proteins that span membranes multiple times may include amino acids with charged, polar side chains, provided that these side chains interact between helices so that they are shielded from the fatty acid environment in the membrane. Because of these hydrophilic interactions, such proteins can create pores for the transport of polar molecules and ions; we will see some of these proteins later. Integral membrane proteins that do not span the membrane also have a hydrophobic helical domain that anchors them in the membrane, while their hydrophilic domains typically interact with intracellular or extracellular molecules to e.g., hold cells in place give cells and tissues their structure, etc.
The very presence of the hydrophobic alpha-helical domains in trans-membrane proteins makes them difficult if not impossible to isolate from membranes in a biologically active form. By contrast, the peripheral polypeptide cytochrome c readily dissociates from the cristal membrane, making it easy to purify. For many years, an inability to purify other cristal membrane electron carriers in biologically active form limited our understanding of the structure and function of the mitochondrial electron transport system.
Hydrophobic alpha-helical domains are in fact, a hallmark of membrane-spanning proteins. It is even possible to determine the primary structure of a polypeptide encoded by a gene before the protein itself has been isolated. For example, knowing the DNA sequence of a gene, we can infer the amino acid sequence of the protein encoded by the gene. A hydrophobicity analysis of the inferred amino acid sequence can tell us if a protein is likely to be a membrane protein. Let’s look at a hydropathy (hydrophobicity) plot (below).
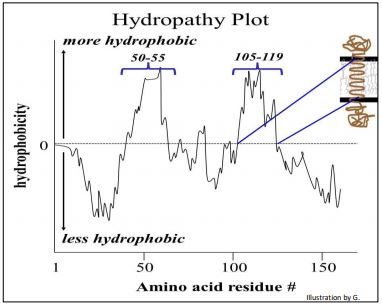
To see how an hydropathy plot can predict whether a protein is a membrane protein, check out the link below.
287 Hydropathy Predicts Hydrophobic Domains and Membrane Proteins