
14.3: The 'Jumping Genes' of Maize
- Page ID
- 16501

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Barbara McClintock’s report that bits of DNA could jump around and integrate themselves into new loci in DNA was so dramatic and arcane that many thought the phenomenon was either a one-off, or not real! Only with the subsequent discovery of transposons in bacteria (and in other eukaryotes) were McClintock’s jumping genes finally recognized for what they were! As we describe her experiments, keep in mind that McClintock’s research and intuitions about gene regulation and epigenetic inheritance came long before molecular technologies made it possible to prove and give names to these phenomena. To begin our tale of transposons, look at the illustration of maize reproduction below.

The different colors of corn seeds (ker nels) result from anthocyanin pigments that are expressed differentially by cells of the aleurone tissue. Mclintock was studying the inheritance of color variation, which ranged from colorless (white or yellow due to an absence of anthocyanins) to brown, purple, spotted or streaked.
The mosaic of kernel colors are vividly shown the corncobs in the photograph below

Clearly, kernel color is inherited. The inheritance of colorless and purple seed color did indeed follow Mendelian rules, but the genetics of mosaicism did not. Mosaic color patterns after genetic crosses were not consistent, implying that the mutations responsible for kernel color were not due to mutations in germ cells. Rather, genes controlling anthocyanin synthesis must be undergoing mutations in somatic cells that would become (or already were) the ones in which the pigments were produced.
242 What Interested McClintock About Maize
A. Discovering the Genes of Mosaicism; the Unstable Ds Gene
McClintock was looking for a genetic explanation for seed color variation in the 1940s and early 1950s. DNA structure had only recently been published. Gene cloning and DNA sequencing were decades into the future! Her only available technologies were based on understanding Mendelian allelic assortment in traditional breeding studies. Nevertheless, since seed color is expressed in cells derived from endosperm, she knew that the inheritance of kernel color phenotype must be studied against a triploid genetic background. McClintock was also aware of proposals that the variegated color phenotype might result when an ‘unstable mutation’ that produced colorless kernels ‘reverted’ in some cells but not others to create a spotted or streaked phenotype. Just what made for an ‘unstable mutation’ was of course, unknown. McClintock ultimately identified three genes involved in seed kernel coloration. Two of the genes initially studied by McClintock controlled the presence vs. absence of kernel color. These are the C and Bz genes:
- C' is the dominant inhibitor allele, so-called because if even one copy was present, the kernels were colorless (yellow), regardless of the rest of the genetic background
- Bz and bz are dominant and recessive alleles of the Bz gene, respectively. In the absence of a C’ (dominant) allele, the presence of a Bz allele would lead to purple kernels. If the bz allele was present without both C’ and Bz alleles, the kernels would be dark brown.
- The gene required to get variegated kernel color was the Ds (Dissociator) gene. Without a viable Ds gene, kernels were either colored or colorless depending on the possible genotypes dictated by the C and Bz alleles.
In other words, it must be the Ds gene that suffers ‘unstable mutations. Because the Ds gene effect was random and only affected some aleurone layer cells, it was suspected to be a region of chromosomal instability (prone to damage or breakage) in some cells but not others. Let’s look at what McClintock did to figure out what was going on in corn kernel color genetics.
Having already demonstrated crossing-over in maize (actually, another remarkable achievement!), McClintock mapped the C’, Bz and Ds genes to Chromosome 9. She then selectively mated corn with the genotypes shown in the protocol below.
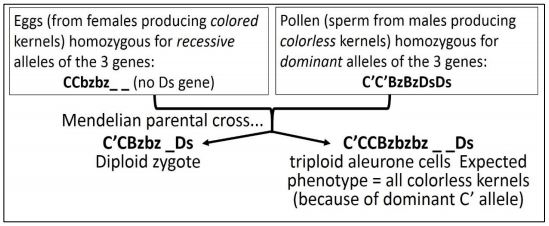
Remember that triploid cell genotypes are being considered in this illustration! You can refer to the phenotypic effects of the allelic backgrounds of three genes as we follow McClintock’s cross. Her cross of a homozygous recessive with a homozygous dominant plant should ring a bell! Let’s look more closely at this cross.
The expected triploid genotypes from the cross are shown below.
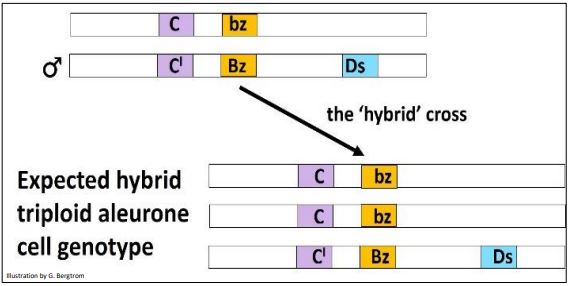
Aleurone cells resulting from this cross should all be colorless (yellow) because of the presence of the dominant C’ allele. However, while there were indeed many colorless kernels on the hybrid cob, there were also many mosaic kernels with dark spots/streaks against a colorless background. McClintock’s interpretation of events is illustrated below
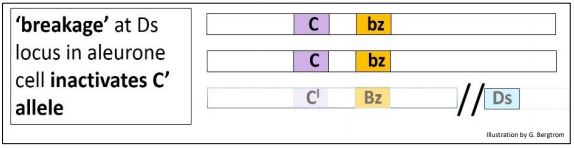
According to McClintock, if some aleurone layer cells in some kernels suffered chromosome breakage at the Ds (Dissociator) locus (indicated by the double slash, //), the C’ allele is inactivated. Without a functional C’ allele, the operative genotype in the affected cells is CCbzbz. These cells then revert to making the brow pigment as directed by the bz allele. When these cells divide, they create clusters of brown cells surrounded by cells with an unbroken chromosome and thus an active C’ allele, creating the appearance of pigment spots or streaks in the kernel, against the otherwise colorless background in the surrounding cells.
243 Variegated Maize Kernels result from "Loss" of the Ds Gene
B. The Discovery of Mobile Genes: the Ac/Ds System
The experiments just described were reproducible using a single breeding stock of maize. But when McClintock tried to repeat the experiments by crossing the homozygous dominant males with homozygous recessive females from a different breeding stock, all the kernels of the progeny cobs were colorless, as if the Ds gene had not caused any chromosomal damage.
It seemed that the Ds gene contributed by the male was unable to function (i.e., ‘break’) in females of this new breeding stock. McClintock hypothesized that the female in the original cross must have contributed a factor that could somehow activate the Ds gene to break, and that this factor, yet another gene, was absent or inactive in the females of the new breeding stock. McClintock called the new factor the activator, or Ac gene. Based on the dependence of Ds on the Ac locus, McClintock recognized that these ‘genes’ were part of as a 2-element, Ac/Ds system influencing mosaicism in maize kernels.
She then demonstrated that Ac-dependent Ds ‘breakage’ was in some cases also associated with inactivation of a normal Bz gene, leading to a loss of purple color kernels. It was at this point that McClintock concluded that far from simply ‘breaking’ the chromosome at a fragile Ds locus, the Ds gene had actually moved to (or into) the Bz gene, disrupting its function. Again, this could not happen in the absence of an active Ac gene. McClintock had discovered the first transposon, earning the 1983 Nobel Prize in Physiology or Medicine! With the advent of recombinant DNA technologies, we now know that:
- The Ds element is a transposon missing a gene for a transposase enzyme required for transposition.
- The Ac element has this gene and is capable of independent transposition.
- Ac provides the transposase needed to mobilize itself and the Ds element
- The sequence similarity of Ds and Ac elements support their common ancestory.
The basic features of the maize Ac/Ds system are:
- Ac is 4563 bp long.
- Ds is a truncated version of Ac.
- There are eleven bp inverted repeats at either end of the Ac and Ds element.
- There are eight bp direct repeats (NOT inverted repeats) of 'target DNA' at the site of insertion of either transposon.
Look for these features as we describe prokaryotic and eukaryotic transposons.