14.4: Transposons Since McClintock
- Page ID
- 16502

Transposons exist everywhere we look in prokaryotes and account for much of eukaryotic repetitive DNA. As such, they can be a large proportion of eukaryotic genomes, including some that no longer even transpose. Transposons were once considered useless or junk DNA, with no obvious function…, or selfish genes with no other purpose than selfreplication. But in light of some new evidence, perhaps not!
As you will see, mechanisms of transposition share many features with DNA replication, recombination and repair, and even viral infection. As you study these mechanisms, keep in the back of your mind that transposition is often triggered by cellular stress.
A. Insertion Sequences (IS Elements)
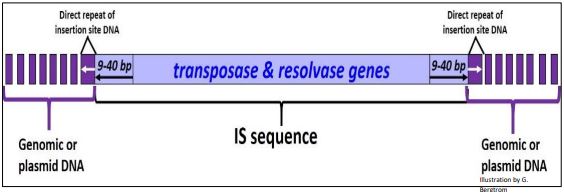
Bacterial IS elements were the first mobile elements described after those in maize. As we’ll see, they share some structural features of eukaryotic transposons. Discovered in the late 1960s, many have been identified (IS1, IS2…, IS10 etc.). Some are inserted into well-known genes (e.g., those of the lac operon), but most are not, likely because there is little ‘extra’ DNA in the compact bacterial genome. Without extra non-coding DNA to buffer against damaging mutations, few bacterial cells would live to tell a tale of transposition! It should surprise us that IS elements can be made to transpose in the lab, but are generally silent in nature.
Members of the IS element family vary in length from about 750 to 1425 bp. Within this stretch of DNA lie transposase and resolvase genes whose products are necessary for mobility. At either end of the IS element are inverted repeats, and when found in either genomic or plasmid DNA, the IS sequence itself is flanked by direct repeats of host genome or plasmid DNA that result from the mechanism of transposition. Again, because of their compact genomes, bacteria can only tolerate low copy numbers of IS elements in their genome or on plasmids (less than ten copies and as few as one!). A typical IS element is illustrated below.
B. Composite Transposons: Tn Elements
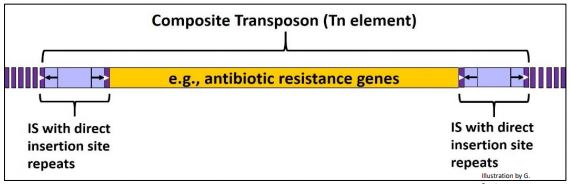
If a pair of IS elements should lie close to each other, separated by a short stretch of genomic or plasmid DNA, they can transpose together, carrying the DNA between them as part of a composite transposon, or Tn element. If some of the DNA between IS elements in a Tn element contains antibiotic resistance genes, its transposition can carry and spread these genes to other DNA in the cell. Tn elements (like IS elements) are present in low copy number. A generic Tn element is drawn below.
Antibiotic resistance genes have the medical community worried; their spread has led to antibiotic-resistant pathogens that cause diseases that are increasingly hard and even impossible to treat. Earlier we saw genetic ‘transformation’ of streptococcal cells that pick up virulence genes in DNA from dead cell. We routinely transform cells with plasmids as part of recombinant DNA experiments. But bacteria can transfer plasmid DNA between themselves quite naturally. During bacterial conjugation, an F (fertility) plasmid normally transfers DNA between compatible bacterial mating types (review bacterial conjugation elsewhere in this text for more details). An F plasmid containing a Tn element harboring an antibiotic resistance gene can thus is passed from donor to recipient during conjugation. The Tn element can transpose into to the recipient bacterial genome. In this way, transposition is a major pathway for the transfer and spread of antibiotic resistance.
C. Complex Transposons
Bacterial Complex Transposons also contain other genes in addition to those required for mobility. Some complex transposons resemble a bacteriophage, or as in the case of phage Mu, actually are phage! In fact, Mu can function either as an infectious phage that reproduces in an infected cell, or as a transposon in the bacterial genome. Transposon genes in Mu phage are illustrated below.

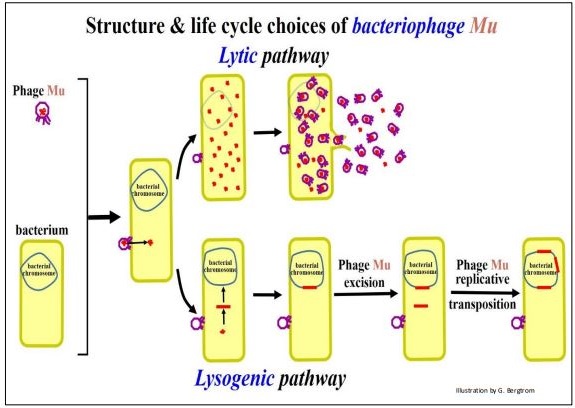
After infecting a bacterium, Mu can enter the lytic phase of its life cycle, replicating its DNA, producing and ultimately releasing new infectious phage ‘particles’ by lysing the host bacterial cell. Alternatively, like other phage, Mu can undergo lysogeny, inserting its DNA into the host cell chromosome. Integrated copies of Mu might excise and re-enter the lytic phase to produce more phage, particularly if some environmental stress threatens host bacterial survival. But, a third lifestyle choice, transposition, is available to Mu once the phage integrates into the bacterial chromosome. The three lifestyle options for Mu phage are illustrated in the next few pages.
Lytic and lysogenic lifestyle options for Mu phage are shown below.

Mu phage DNA can act as a transposable element while in the lysogenic pathway, as shown below
As we turn to a description of eukaryotic transposons, look for similarities to bacterial IS and Tn elements.
D. Overview of Eukaryotic Transposable Elements
There are two classes transposons in eukaryotes:
Class I (Retrotransposons) move/’jump’ by transcription of RNA at one locus, followed by reverse transcription and integration of the cDNA back into genomic DNA at a different location. Retrotransposons may be derived from (or be the source of) retroviruses since active retroviruses excise from and integrate into DNA much like retrotransposons. Retroposons are a sub-class of retrotransposons (see below).
Class II (DNA Transposons) move by one of two mechanisms. In the cut-&-paste pathway, the transposon leaves one locus and integrates at another. In the replicative pathway, the original transposon remains in place while new copies are mobile. The table below shows the distribution and proportion of genomes represented by different classes/types of transposable elements.

The table confirms that bacteria contain few transposons, while eukaryotes vary widely in transposon load (transposons as a percentage of genomic DNA), from as low as 4% to more than 70%.
The table below summarizes transposable elements by class, sub-type, size, genomic distribution, mechanism of transposition, etc.

Between the two tables above, we can conclude the following:
- Transposon load is not correlated with evolutionary complexity of organisms.
- Shared Transposons have different evolutionary histories in different organisms.
- Where transposons remain active, they continue to shape genomic landscapes, especially in organisms with a high transposons load.
We will revisit some of these conclusions later, after looking at the structure and mechanism of mobility of different transposable elements.
E. The Structure of Eukaryotic DNA (Class II) Transposons
Active eukaryotic DNA transposons share structural features with bacterial mobile elements, including genes required for transposition, flanking inverted repeats and flanking insertion-site direct repeats of host cell DNA. The characteristic structure of a eukaryotic DNA transposon is shown below.
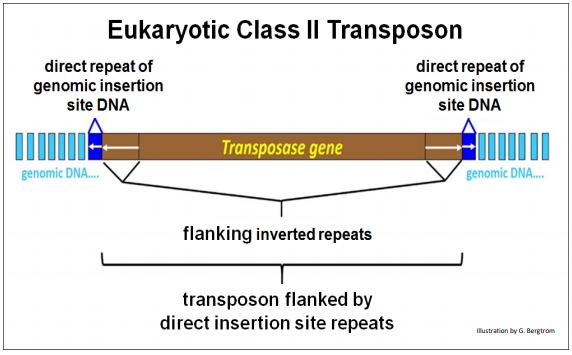
Class II transposons can ‘jump’ by cut-and-paste or replicative mechanisms. Cut-and-paste transposition removes a copy from one location and moves (transposes) it to another location. As its name suggests, replicative transposition leaves a copy of the original transposon in place while inserting a new copy elsewhere in the genome. Transposition by the cut-and-paste mechanism is diagrammed below.

Note that after transcription of the transposase gene, the enzyme nicks the DNA and trims the 3’OH ends to create a staggered cut to excise the transposon. The transposase actually brings the transposon ends together during the cut step and mediates its insertion at a new DNA site. After ligation of the 3’OH ends of the transposon to the 5’OH at the insertion site, replication replaces the missing bases, generating the direct repeats of host cell genomic DNA at the insertion site. A final ligation step completes transposition.
In the replicative transposition, the transposon also nicks and trims the DNA at its source (original) insertion site. But, unlike the cut and paste mechanism, the source transposon is not excised.
Details of the replicative mechanism of transposition is summarized below.

After nicking the 3’ ends of the transposon at the insertion site, transposase holds the transposon ends together while catalyzing a hydrolytic attack of DNA at a new insertion site. This is followed by priming of transposon strand replication from the 3’OH ends of the insertion site DNA strands. A cointegrate structure forms in which each transposon copy has been made by semi-conservative replication. The cointegrate is resolved by one of two recombinational mechanisms. The result leaves copies of the transposon at both the original site and the new insertion site.
Let’s compare and contrast the features of cut-&-paste and replicative DNA transposition. The common features are that:
- Transposon-encoded transposase binds, brings transposon ends together and catalyzes single-stranded cleavage (hydrolysis) leaving ‘staggered ends’.
- Transposase holds the transposon ends together for the remaining steps.
The differences between the two mechanisms are that in cut & paste transposition, the transposon is completely excised and then transposed to a new site in genomic DNA. In contrast, after single stranded cleavage in replicative transposition, transposase-bound free 3’ ends of the transposon hydrolyze both strands of stranded DNA at a new insertion site. After ligation of the 3’ ends of transposon strands to 5’ ends of cut genomic DNA insertion-site ends, the remaining 3’ ends of the insertion site DNA ends prime replication of the transposon, forming the cointegrate, which is followed by its resolution by one of two recombination pathways.
F. The Structures of Eukaryotic RNA (Class I) Transposons
Like DNA transposons, all RNA transposons leave insert-site footprints, i.e., direct repeats of genomic DNA flanking the element. Unlike DNA transposons, active eukaryotic Class I transposons move via an RNA intermediate. Also unlike DNA transposons, they lack terminal inverted repeats.
The mobility of the RNA intermediate of all retrotransposons requires a promoter that recognizes a reverse transcriptase enzyme as well as endonuclease and integrase enzymes (to be described below). Autonomous Class I RNA transposons include LTR retrotransposons (e.g., the yeast Ty element) as well as Non-LTR retrotransposons). The latter include the autonomous LINEs (Long Interspersed Nuclear Elements). The autonomous LTR and Non-LTR LINEs contain and express genes needed for enzymes required for transposition. On the other hand, SINEs (a sub-class of Non-LTR retrotransposons) lack genes for enzymes required for transposition and therefore can’t transpose independently. Thus, they are non-autonomous retrotransposons that rely on “true” (autonomous) retrotransposon activity for mobility. SINEs are sometimes called retroposons to distinguish them from the autonomous retrotransposons.
249 Introduction to Features of Retrotransposition
Next we take a closer look at Retrotransposon structures and the genes and enzyme activities required for retrotransposition
1. LTR retrotransposons: The Yeast Ty Element
The Ty transposon harbors several genes needed for transposition. These include:
- the Gag gene that encodes group-specific antigen, a protein that forms a viruslike particle that will contain reverse-transcribed transposon DNA,
- the RT gene that encodes the reverse transcriptase that will make reversetranscribed copies of retrotransposon transcript RNAs.
- the Prt gene that encodes a protease that will break down the virus-like particle as the retrotransposon enters the nucleus.
- the Int gene that encodes the integrase required for integration of the retrotransposon into a genomic DNA insertion site.
A representative Ty element is shown below as it would exist integrated into yeast genomic DNA.
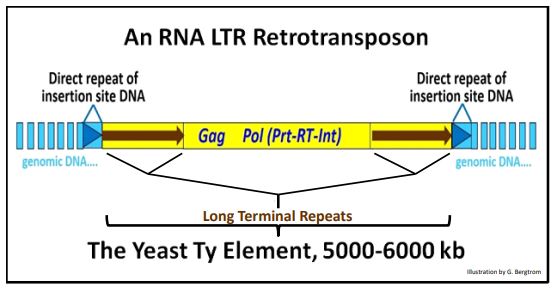
In fact, many of the events in Ty transposition occur in the cytoplasmic “virus-like particle” in yeast cells. To see more, click here. Note that the Pol region in the illustration above consists of overlapping open reading frames (ORFs) encoding the Prt, RT and Int genes. The ready-to-move transposon consists only of the region of DNA symbolized in yellow.
2. Non-LTR Retrotransposons: LINEs
LINEs (Long Interspersed Nuclear Elements) also encode enzymes needed for transposition and like other transposons, generate target-site direct repeats flanking the inserted element. But they do not have the long terminal repeats! Instead, their ORFs (genes) are flanked by 5’ and a 3’ untranslated regions (UTRs).
The structure of the human L1 Line is drawn below.
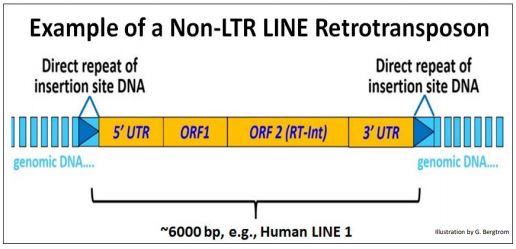
The 5’ UTR contains a promoter from which cellular RNA polymerase II can transcribe the downstream genes (see the Transcription chapter). The second of these (ORF2) encodes the reverse transcriptase and an integrase activity essential for transposition of the LINE. All Class I (RNA-intermediate) autonomous transposons share the following features:
a) a Promoter in the 5’ UTR from which they can be transcribed.
b) a Reverse Transcriptase that generates a cDNA copy of the transposable element.
c) RNAse H (an endonuclease) that degrades that transcript after reverse transcription.
d) Integrase (like a transposase) that catalyzes insertion of the retrotransposon copy at insertion sites.
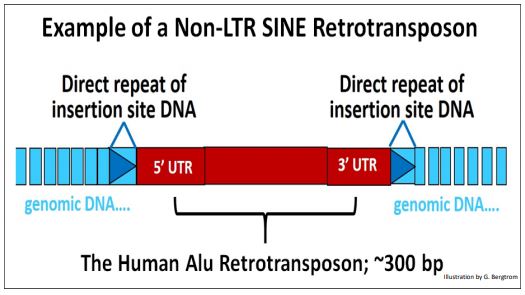
3. Non-LTR SINE retrotransposons
Non-LTR SINE retrotransposons typically lack genes, but their non-genic DNA is nonetheless flanked by 5’ and 3’ UTRs. RNA polymerase III, which also transcribes transfer RNAs, also transcribes SINEs. However, to transpose, they rely on the concurrent activity of a Non-LTR transposon (a LINE) to provide the requisite enzymatic activities.
A typical SINE (e.g., the Alu element) is shown below.
G. Mechanisms of Retrotransposition
There are two mechanisms of retrotransposition: Extrachromosomally Primed Retrotransposition (LTR retrotransposons for example) and Insertion Target-Site Primed Retrotransposition (non-LTR Retrotransposons like LINEs and SINEs). These will be considered next.
1. Extrachromosomally Primed Retrotransposition (e.g., of a LINE)
As its name suggests, in extrachromosomally primed retrotransposition, a circular reverse transcript of the retrotransposon attacks, nicks and integrates into a genomic insertion site. In this mechanism, reverse transcriptase creates a cDNA copy of a transcribed retro-element. Integrase/endonuclease then binds the cDNA copy, holding the ends together, in effect circularizing it. This isolable ribonucleoprotein resembles an intasome, a structure similar to the nucleoprotein complex that catalyzes the integration of retroviral cDNAs during lysogeny.
Extrachromosomally primed retrotransposition is illustrated below.
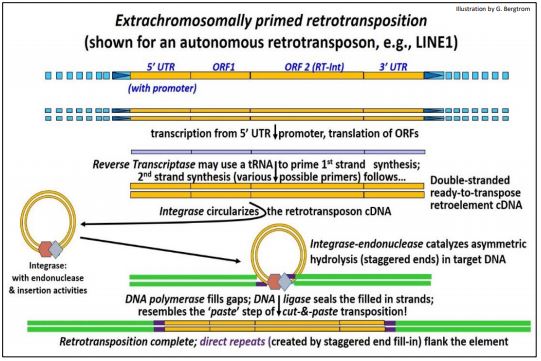
The three-dimensional structure of a retroviral intasome interacting with DNA and nucleosomes was recently determined (for more, see Retroviral Intasome 3D Structure). In this form, the retrotransposon attacks DNA at an insertion site, creating staggered ends. After insertion, the gaps in the DNA are filled in. Ligation seals the retrotransposon in its new location, creating direct insertion site repeats.
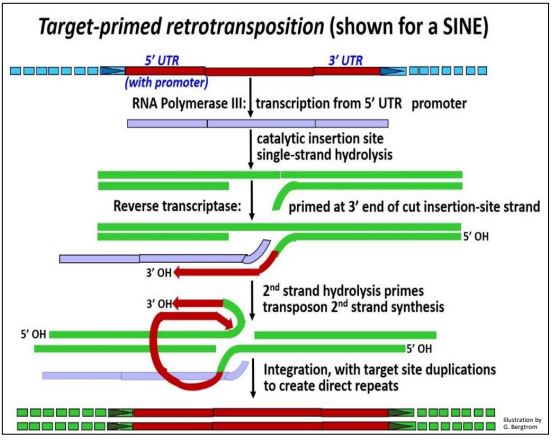
2. Target-Site Primed SINE Retrotransposition (e.g., of a SINE)
A key feature of target-site primed retrotransposition (retroposition) is the absence of an integrase-bound, circular double-stranded reverse transcript. In SINE transposition, RNA polymerase III (the same enzyme that catalyzes tRNA and 5S rRNA transcription) transcribes the SINE. If a LINE is concurrently transcribed, its enzymes will be made. When its integrase-endonuclease catalyzes hydrolysis of one strand of DNA at a new insertion site, the 3’OH end of this strand can prime reverse transcription of the one SINE cDNA strand by the LINE reverse transcriptase. After hydrolysis of the second target site DNA strand, its 3’-OH end primes replication of the second strand of the SINE cDNA. Integrase completes insertion of the copy-SINE in its new genomic location. The target-site primed retrotransposition mechanism of retrotransposition is illustrated below.