
12.4: Gene Regulation in Eukaryotes
- Page ID
- 16488

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)A. The Difference between Eukaryotic and Prokaryotic Gene Regulation
Let's Recall an experiment described earlier and illustrated below.
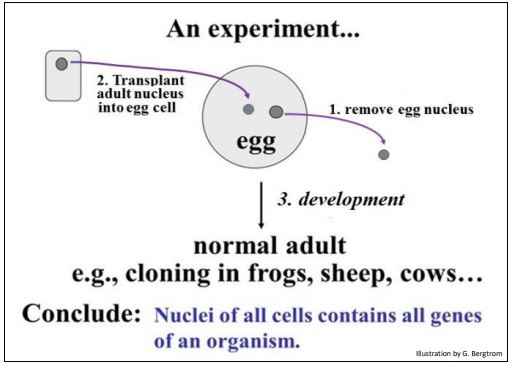
Results of this experiment provided the evidence that even very different cells of an organism contain the same genes. In fact, in any multicellular eukaryotic organism, every cell contains the same DNA (genes). Therefore, the different cell types in an organism differ not in which genes they contain, but which sets of genes they express! Looked at another way, cells differentiate when they turn on new genes and turn off old ones. Thus, gene regulation produces different sets of gene products during differentiation, leading to cells that look and function differently in the organism.
220 An Experiment: All of an Organism's Cells Have the Same Genome
Compared to prokaryotes, many steps in eukaryotes lie between transcription of an mRNA and the accumulation of a polypeptide end product. Eleven of these steps are shown in the pathway from gene to protein below.
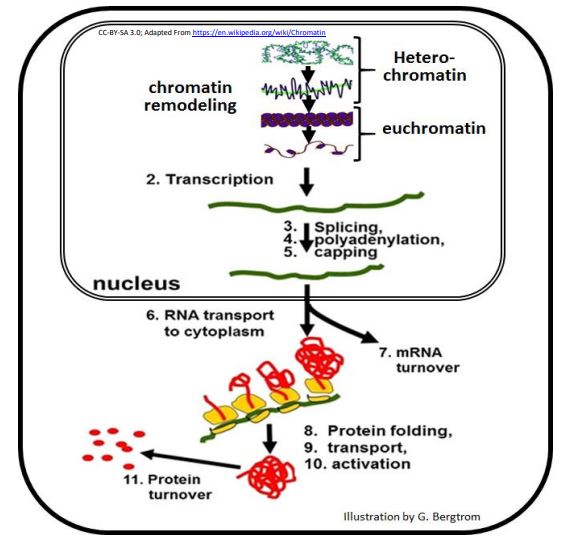
Theoretically, cells could turn on, turn off, speed up or slow down any of the steps in this pathway, changing the steady state concentration of a polypeptide in the cells. While regulation of any of these steps is possible, the expression of a single gene is typically controlled at only one or a few steps. A common form of gene regulation is at the level of transcription initiation, similar to transcriptional control in bacteria, in principle if not in detail.
B. Complexities of Eukaryotic Gene Regulation
Gene regulation in eukaryotes is more complex than in prokaryotes. This is in part because their genomes are larger and because they encode more genes. For example, the E. coli genome houses about 5,000 genes, compared to around 25,000 genes in humans. Furthermore, eukaryotes can produce even more than 25,000 proteins by alternative splicing of mRNAs and in at least a few cases, by initiating transcription from alternative start sites in the same gene. And of course, the activity of many more genes must be coordinated without the benefit of multigene operons! Finally, eukaryotic gene regulation is made more complicated because all nuclear DNA is wrapped in protein in the form of chromatin.
All organisms control gene activity with transcription factors that bind to specific DNA sequences (cis regulatory elements). In eukaryotes, these elements can be proximal to (near) the promoter of a gene, or distal to (quite far from) the gene they regulate. A eukaryotic map showing the components of a typical gene and its associated cis-acting regulatory elements is shown below.
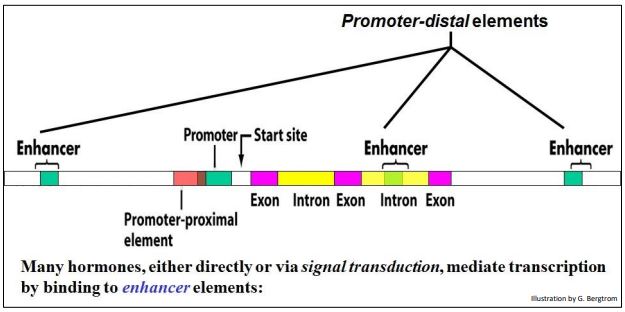
Enhancers are typical distal cis elements that recognize and bind transcription factors to increase the rate of transcription of a gene. Oddly enough, these short DNA elements can be in the 5’ or 3’ non-translated region of the gene, or even within introns, and can lie thousands of base pairs away from the genes they control. Note that enhancer elements are even in introns can also be very far from the start-site of transcription of a gene.
Upstream regulatory regions of eukaryotic genes (to the left of a gene promoter as shown above) often have distal binding sites for more than a few transcription factors, some with positive (enhancing) and others with negative (silencing) effects. Of course, which of these DNA regions are active in controlling a gene depends on which transcription factor(s) are present in the nucleus. Sets of positive regulators will work together to coordinate and maximize gene expression when needed, and sets of negative regulators will bind negative regulatory elements to silence a gene.
222 Transcription Factors Bind DNA Near and Far
We saw that in eukaryotes, the initiation of transcription involves many transcription factors and RNA polymerase II acting at a gene promoter to form a transcription preinitiation complex. TFIID, or TATA binding protein is one of the first factors to bend, causing the DNA in the promoter region to bend, much like the CAP protein in bacteria. TFIID also recruits other transcription factors to the promoter. As in bacteria, bending the DNA loosens H-bonds between bases, facilitating unwinding the double helix near the gene. Bending eukaryotic DNA also brings distal regulatory proteins bound to enhancer sequences far from the promoter together with the proteins bound to more proximal regulatory elements, as shown in the drawing below.
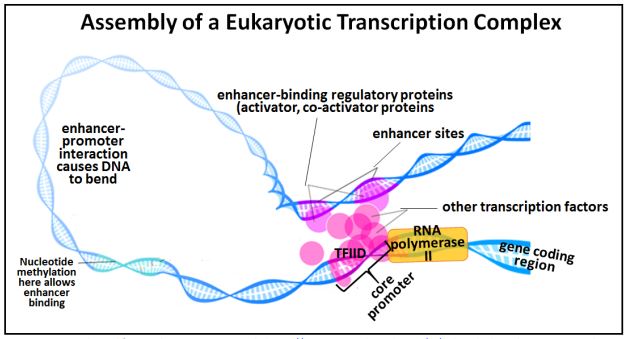
Nucleotide methyation sites may facilitate regulatory protein-enhancer binding. When such regulatory proteins, here called activators (i.e., of transcription), bind to their enhancers, they acquire an affinity for protein cofactors that enable recognition and binding to other proteins in the transcription initiation complex. This attraction stabilizes the bend in the DNA that then makes it easier for RNA polymerase II to initiate transcription
223 Assembling a Eukaryotic Transcription Initiation Complex
It is worth reminding ourselves that it is shape and allosteric change that allow DNAprotein interactions (in fact, any interactions of macromolecules). The lac repressor we saw earlier is a transcription factor with helix-turn-helix DNA binding motifs. This motif and two others (zinc finger, and leucine zipper) characterize DNA binding proteins are illustrated below.
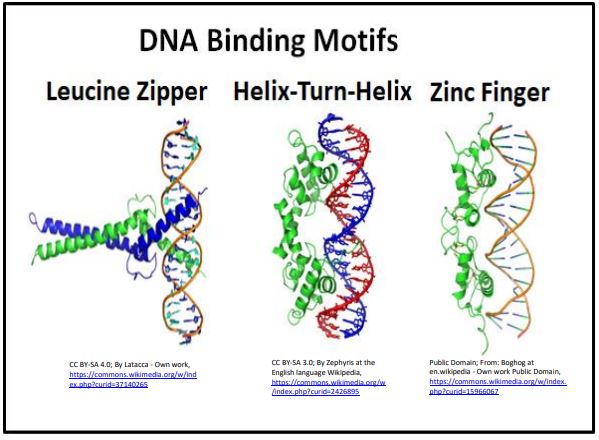
DNA-binding motifs in each regulatory protein shown here bind one or more regulatory elements ‘visible’ to the transcription factor in the major groove of the double helix.
224 Transcription Factor Domains/Motifs Bind Specific DNA Sequences
We will look next at some common ways in which eukaryotic cells are signaled to turn genes on or off, or to increase or decrease their rates of transcription. As we describe these models, remember that eukaryotic cells regulate gene expression in response to changes in extracellular environments. These can be unscheduled, unpredictable changes in blood or extracellular fluid composition (ions, small metabolites), or dictated by changes in a long-term genetic program of differentiation and development. Changes in gene expression even obey circadian (daily) rhythms, the ticking of a clock. In eukaryotes, changes in gene expression, expected or not, are usually mediated by the timely release of chemical signals from specialized cells (e.g., hormones, cytokines, growth factors, etc.). We will focus on some betterunderstood models of gene regulation by these chemical signals.
C. Regulation of Gene Expression by Hormones that enter Cells and Those That Don't
Gene-regulatory (cis) elements in DNA and the transcription factors that bind to them have co-evolved. But not only that! Eukaryotic organisms have evolved complete pathways that respond to environmental or programmed developmental cues and lead to an appropriate cellular response. Chemicals that regulate genes in prokaryotes are not usually signals communicated by other cells. In eukaryotes, chemicals released by some cells signal other cells to respond, thus coordinating the activity of the whole organism. Hormones released by cells in endocrine glands are well-understood signal molecules; hormones affect target cells elsewhere in the body.
225 Chemicals That Control Gene Expression
1. How Steroid Hormones Regulate Transcription
Steroid hormones cross the cell membranes to have their effects. Common steroid hormones include testosterone, estrogens, progesterone, glucocorticoids and mineral corticoids. Once in target cell, such hormones bind to a steroid hormone receptor protein to form a steroid hormone-receptor complex. The receptor may be in the cytoplasm or in the nucleus, but in the end, the hormone-receptor complex must bind to DNA regulatory elements of a gene to either enhance or silence transcription. Therefore, a steroid hormone must cross the plasma membrane, and may also need to cross the nuclear envelope.
Follow the Binding of a steroid hormone to a cytoplasmic receptor below.
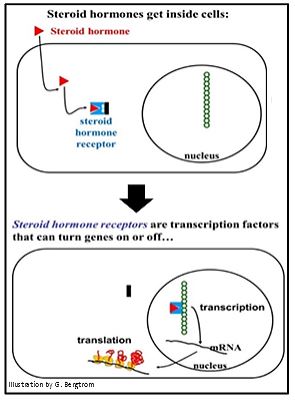
Here the hormone (the triangle) enters the cell. An allosteric change in the receptor releases a protein subunit called Hsp90 (the black rectangle in the illustration). The remaining hormone-bound receptor enters the nucleus.
The fascinating thing about Hsp90 is that it was first discovered in cells subjected to heat stress. When the temperature gets high enough, cells shut down most transcription and instead transcribe Hsp 90 and/or other special heat shock genes. The resulting heat shock proteins seem to protect the cells against metabolic damage until temperatures return to normal. Since most cells never experience such high temperatures, the evolutionary significance of this protective mechanism is unclear. As we now know, heat shock proteins have critical cellular functions, in this case blocking the DNA-binding site of a hormone receptor until a specific steroid hormone binds to it.
Back to hormone action! No longer associated with the Hsp90 protein, the receptor bound to its hormone cofactor binds to a cis-acting transcription control element in the DNA, turning transcription of a gene on or off. The hormone receptors for some steroid hormones are already in the nucleus of the cell, so the hormone must cross not only the plasma membrane, but also the nuclear envelope in order to access the receptor.
As for steroid hormone functions, we already saw that glucocorticoids turn on the genes of gluconeogenesis. Steroid hormones also control sexual development and reproductive cycling in females, salt and mineral homeostasis in the blood, metamorphosis in arthropods, etc., all by regulating gene expression.
2. How Protein Hormones Regulate Transcription
Protein hormones are of course large and soluble, with highly charged surfaces. Other hormones might be relatively small (e.g., adrenalin), but are charged. Large or highly charged signal molecules cannot get across the phospholipid barrier of the plasma membrane. To have any effect at all, they must bind to receptors on the surface of cells. These receptors are typically membrane glycoproteins.
The information (signals) carried by protein hormones must be conveyed into the cell indirectly, by a process called signal transduction. There are two well-known pathways of signal transduction, each of which involves activating pathways of protein phosphorylation in cytoplasm. The phosphorylation cascade that results activates a transcription factor that binds to regulatory DNA, turning a gene on or off.
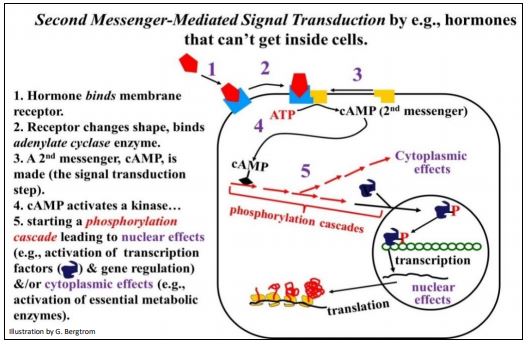
Binding of a hormone to a cell surface receptor leads to an allosteric change in the receptor. This in turn activates other proteins either in the plasma membrane or in the cytoplasm, leading to the synthesis of a cytoplasmic second messenger. The second messenger typically binds to a protein kinase in the cytoplasm, launching a series of protein phosphorylations, or a phosphorylation cascade. The last in the series of proteins to be phosphorylated is an activated transcription factor that will bind to a cis-regulatory DNA sequence.
cAMP was the first second messenger metabolite to be discovered. It mediates many hormonal responses, controlling both gene activity and enzyme activity. cAMP forms when the hormone-receptor in the membrane binds to and activates a membrane-bound adenylate cyclase enzyme. The cAMP produced then binds to a protein kinase, the first of several in a phosphorylation cascade. Signal transduction mediated by cAMP is summarized in the illustration below.
A different kind of signal transduction involves a hormone receptor that is itself the protein kinase. The role of enzyme-linked hormone receptors in signal transduction is summarized below.
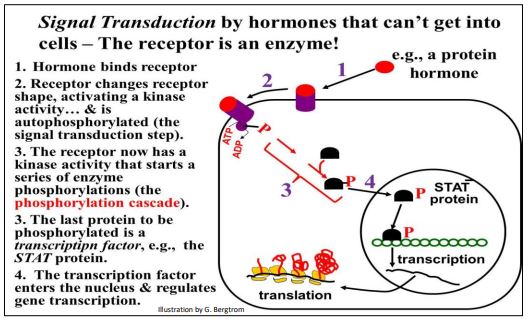
Binding of the signal protein (e.g. hormone) to the enzyme-linked receptor causes an allosteric change that activates the receptor kinase, starting phosphorylation cascade resulting in an active transcription factor. We look at signal transduction in more detail in another chapter.
D. Regulating Eukaryotic Genes Means Contending with Chromatin
Consider again the illustration of the different levels of chromatin structure (below).
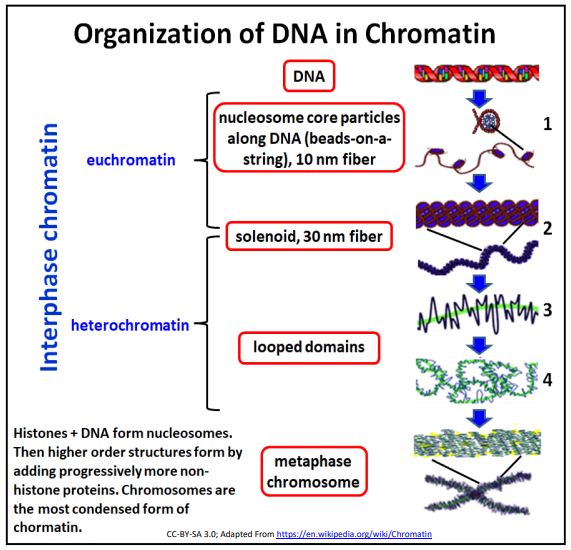
Transcription factors bind specific DNA sequences by detecting them through the grooves (mainly the major groove) in the double helix. The drawing above reminds us however, that unlike the nearly naked DNA of bacteria, eukaryotic (nuclear) DNA is coated with proteins that, in aggregate are by mass, greater than the mass of DNA that they cover. The protein-DNA complex of the genome is of course, chromatin.
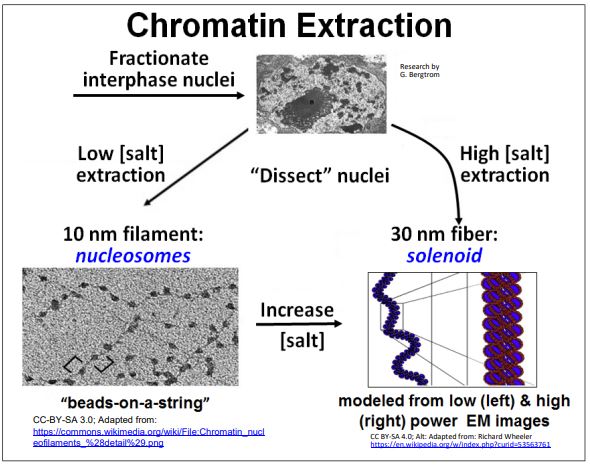
Again, as a reminder, DNA coated with histone proteins forms the 9 nm diameter beads-on-a-string structure in which the beads are the nucleosomes. The association of specific non-histone proteins causes the nucleosomes to fold over on themselves to form the 30 nm solenoid. As we saw earlier, it is possible to selectively extract chromatin. Take a second look at the results of typical extractions of chromatin from isolated nuclei below.
Further accretion of non-histone proteins leads to more folding and the formation of euchromatin and heterochromatin characteristic of non-dividing cells. In dividing cells, the chromatin further condenses to form the chromosomes that separate during either mitosis or meiosis.
Recall that biochemical analysis of the 10 nm filament extract revealed that the DNA wraps around histone protein octamers, the nucleosomes or beads in this beads-on-astring structure. Histone proteins are highly conserved in the eukaryotic evolution (they are not found in prokaryotes). They are also very basic (many lysine and arginine residues) and therefore very positively charged. This explains why they are able to arrange themselves uniformly along DNA, binding to the negatively charged phosphodiester backbone of DNA in the double helix.
Since the DNA in euchromatin is less tightly packed than it is in heterochromatin, perhaps active genes are to be found in euchromatin and not in heterochromatin. Experiments in which total nuclear chromatin extracts were isolated and treated with the enzyme deoxyribonuclease (DNAse) revealed that the DNA in active genes was degraded more rapidly than non-transcribed DNA. More detail on these experiments can be found in the two links below.
228 Question: Is Euchromatic DNA Transcribed?
229 Experiment and Answer: Euchromatin is Transcribed
The results of such experiments are consistent with the suggestion that active genes are more accessible to DNAse because they are in less coiled, or less condensed chromatin. DNA in more condensed chromatin is surrounded by more proteins, and thus is less accessible to, and protected from DNAse attack. When packed up in chromosomes during mitosis or meiosis, all genes are largely inactive.
Regulating gene transcription must occur in non-dividing cells or during the interphase of cells, where changing the shape of chromatin (chromatin remodeling) in order to silence some and activate other genes is possible. Changing chromatin conformation involves chemical modification of chromatin proteins and DNA. For example, chromatin can be modified by histone acetylation, de-acetylation, methylation and phosphorylation, reactions catalyzed by histone acetyltransferases (HAT enzymes), de-acetylases, methyl transferases and kinases, respectively. For example, acetylation of lysines near the amino end of histones H2B and H4 tends to unwind nucleosomes and open the underlying DNA for transcription. De-acetylation then, promotes condensation of the chromatin in the affected regions of DNA. Likewise, methylation of lysines or arginines (the basic amino acids that characterize histones!) of H3 and H4 can open DNA for transcription, while demethylation has the opposite effect. In one case, di-methylation of a lysine in H3 can suppress transcription. These chemical modifications affect recruitment of other proteins that alter chromatin conformation and ultimately activate or block transcription.
This reversible and its effect on chromatin are illustrated below.
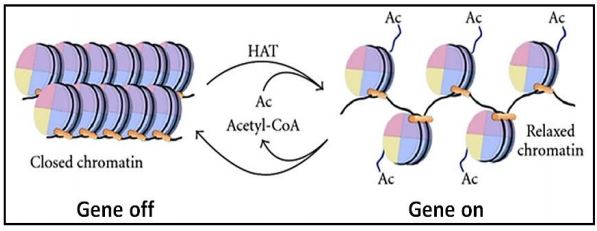
Nucleosomes themselves can be moved, slid and otherwise repositioned by complexes that hydrolyze ATP for energy to accomplish the physical shifts. Some cancers are associated with mutations in genes for proteins involved in chromatin remodeling. This is no doubt, because failures of normal remodeling could adversely affect normal cell cycling and normal replication. In fact, a single, specific pattern of methylation may mark DNA in multiple cancer types (check out Five Cancers with the Same Genomic Signature - Implications).
E. Regulating all the Genes on a Chromosome at Once
Recall that X chromosomes in human female somatic cells is inactivated, visible in the nucleus as a Barr body. One of the two X chromosomes in female fruit flies is also inactivates. However, both males and females of Drosophila (presumably also us!) require X chromosome gene expression during embryogenesis. Given the difference in X chromosome gene dosage between males and females, do males get by with fewer X chromosome gene products than females?
Experiments looking at the expression of X chromosome gene in male and female flies revealed similar levels of gene products. It turns out that the activity of a nuclear body called HLB (Histone Locus Body) is required for increase in X chromosome gene transcription. A protein, called CLAMP (Chromatin-Linked Adaptor for Male-specific lethal (MSL) Protein), was shown to bind to GAGA nucleotide repeats lying between the genes for histones 3 and 4. As there are about 100 repeats of the fivegene histone locus on X chromosomes, and thus about 100 repeats of the GAGA repeats. Therefore, many CLAMP proteins bind to the HLBs, where they recruit many MSL proteins. The MSL protein complexes that form then globally increase male X chromosome gene expression, compensating for the lower X gene dosage in males. Read the original research here (L.E. Reider et al. (2018) Genes & Development 31:1-15). And finally, there is emerging evidence that the HLB action may also be involved in inactivation of an entire female X chromosome later in embryogenesis in females!