
1.3: Domains of Life
- Page ID
- 16410

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)We believe with good reason that all life on earth evolved from a common ancestral cell that existed soon after the origins of life on our planet. Too long ago, not all life was divided into two groups: the true bacteria and everything else!
Now we group life into one of three domains:
They are among the first descendants of that common ancestral cell. They lack nuclei (pro meaning before and karyon meaning kernel, or nucleus). They include bacteria and cyanobacteria (blue-green algae).
Include all higher life forms, characterized by cells with true nuclei (Eu, true; karyon, nucleus)
(meaning “old” bacteria) include many extremophile bacteria (‘lovers’ of life at extreme high temperatures, salinity, etc.). Originally classified as ancient prokaryotes, Archaebacteria were shown by 1990 to be separate from prokaryotes and eukaryotes, a third domain of life.
The archaea are found in such inhospitable environments as boiling hot springs or arctic ice, although some also live in conditions that are more temperate. Carl Woese compared the DNA sequences of genes for ribosomal RNAs in normal bacteria and extremophiles. Based on sequence similarities and differences, he concluded that the latter are in fact a domain separate from the rest of the bacteria as well as from eukaryotes. For a review, see (Woese, C. 2004; A new biology for a new century. Microbiol. Mol. Biol. Rev. 68:173-186)
The three domains of life (Archaea, Eubacteria and Eukarya) quickly supplanted the older division of living things into Five Kingdoms, the Monera (prokaryotes), Protista, Fungi, Plants, and Animals (all eukaryotes!). In a final surprise, the sequences of archaebacterial genes clearly indicate a common ancestry of archaea and eukarya.
Thus, Archaea are not true bacteria! They share genes and proteins as well as metabolic pathways found in eukaryotes but not in bacteria, supporting their close evolutionary relationship to eukaryotes. That they also contain genes and proteins as well as metabolic pathways unique to the group is further testimony to their domain status. Understanding that all living organisms belong to one of three domains has dramatically changing our understanding of evolution. The evolution of the three domains is illustrated below.

A. The Prokaryotes (Eubacteria = Bacteria and Cyanobacteria)
Prokaryotic cells lack a nucleus and other organelles such as mitochondria, chloroplasts, endoplasmic reticulum, and assorted eukaryotic vesicles and internal membranes. Bacteria do contain bacterial microcompartments (BMCs), but these are made up entirely of protein and are not surrounded by a phospholipid membrane.
These function for example in CO2 fixation to sequester metabolites toxic to the cells. Click Bacterial Organelles for more information. Bacteria are typically unicellular, although a few (like some cyanobacteria) live colonial lives at least some of the time. Transmission and scanning electron micrographs of rod-shaped bacteria are shown in the example below at the left. A diagram of bacterial structure is also shown (right).

1. Bacterial Reproduction
Without the compartments afforded by the internal membrane systems common to eukaryotic cells, intracellular chemistries, from DNA replication, transcription, translation, and all the metabolic biochemistry of life, happen in the cytoplasm of the cell. DNA is a circular double helix that duplicates as the cell grows. While not enclosed in a nucleus, bacterial DNA is concentrated in a region of the cell called the nucleoid. When not crowded at high density, bacteria replicate their DNA throughout the life of the cell, dividing by binary fission. The result is the equal partition of duplicated bacterial “chromosomes” into new cells. The bacterial chromosome is essentially naked DNA, unassociated with proteins.
2. Cell Motility and the Possibility of a Cytoskeleton
Movement of bacteria is typically by chemotaxis, a response to environmental chemicals. Some may respond to other stimuli such as light (phototaxy). They can move to or away from nutrients, noxious/toxic substances, light, etc., and achieve motility in several ways. For example, many move using flagella made up largely of the protein flagellin. Flagellin is absent from eukaryotic cells. On the other hand, the cytoplasm of eukaryotic cells is organized by a complex cytoskeleton of rods and tubes made of actin and tubulin proteins. Prokaryotes were long thought to lack these or similar cytoskeletal components. However, two bacterial genes that encode proteins homologous to eukaryotic actin and tubulin were recently discovered. The MreB protein forms a cortical ring in bacteria undergoing binary fission, similar to the actin cortical ring that pinches dividing eukaryotic cells during cytokinesis (the actual division of a single cell into two smaller daughter cells). This is modeled below in the cross-section (right) near the middle of a dividing bacterium (left).
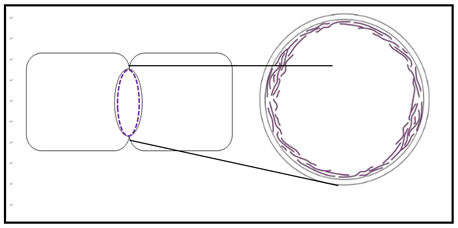
The FtsZ gene encodes a homolog of tubulin proteins. It seems that together with flagellin, the MreB and FtsZ proteins may be part of a primitive prokaryotic cytoskeleton involved in cell structure and motility.
3. Some Bacteria Have Internal Membranes
While bacteria lack organelles (the membrane-bound structures of eukaryotic cells), internal membranes in some bacteria form as inward extensions (invaginations) of plasma membrane. Some of these capture energy from sunlight (photosynthesis) or from inorganic molecules (chemolithotrophy). Carboxysomes are membrane bound photosynthetic vesicles in which CO2 is fixed (reduced) in cyanobacteria (shown below).
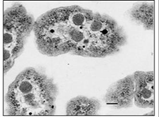
CC-BY; From: en.Wikipedia.org/wiki/File:Carboxysomes_EM.jpg
Photosynthetic bacteria have less elaborate internal membrane systems.
4. Bacterial Ribosomes Do the Same Thing as Eukaryotic Ribosomes… and Look Like Them!
Ribosomes are the protein synthesizing machines of life. Ribosomes of prokaryotes are smaller than those of eukaryotes, but are able to translate eukaryotic messenger RNA (mRNA) in vitro. Underlying this common basic function is the fact that the ribosomal RNAs of all species share base sequence and structural similarities indicating a long evolutionary relationship. Recall similarities revealed the closer relationship of archaea to eukarya than prokarya.
Clearly, the prokarya (Eubacteria) are a diverse group of organisms, occupying almost every wet, dry, hot or cold nook and cranny of our planet. Despite this diversity, all prokaryotic cells share many structural and functional metabolic properties with each other… and with the archaea and eukaryotes! As we have seen with ribosomes, shared structural and functional properties support the common ancestry of all life. Finally, we not only share common ancestry with prokaryotes, we even share living arrangements with them. Our gut bacteria represent up to 10X more cells than our own! Read more at The NIH Human Microbiome Project. Also check out the following link for A Relationship Between Microbiomes, Diet and Disease.
B. The Archaebacteria (Archaea)
Allessandro Volta, a physicist who gave his name to the Volt, discovered methane producing bacteria (methanogens) way back in 1776! He found them living in the extreme environment at the bottom of Lago Maggiore, a lake shared by Italy and Switzerland. These unusual bacteria are cheomoautotrophs that get energy from H2 and CO2 and also generate methane gas in the process. It was not until the 1960s that Thomas Brock (from the University of Wisconsin-Madison) discovered thermophilic bacteria living at temperatures approaching 100oC in Yellowstone National Park in Wyoming. Organisms living in any extreme environment were soon nicknamed extremophiles. One of the thermophilic bacteria, now called Thermus aquaticus, became the source of Taq polymerase, the heat-stable DNA polymerase that made the polymerase chain reaction (PCR) a household name in labs around the world!
Extremophile and “normal” bacteria are similar in size and shape(s) and lack nuclei. This initially suggested that most extremophiles were prokaryotes. But as Carl Woese demonstrated, it is the archaea and eukarya that share a more recent common ancestry! While some bacteria and eukaryotes can live in extreme environments, the archaea include the most diverse extremophiles. Here are some examples of extremophiles:
• Acidophiles: grow at acidic (low) pH.
• Alkaliphiles: grow at high pH.
• Halophiles: require high salt concentrations; see Halobacterium salinarium below.

• Methanogens: produce methane; a cross section of Methanosarcina acetivorans is shown above (right). Note the absence of significant internal structure.
• Barophiles: grow best at high hydrostatic pressure.
• Psychrophiles: grow best at temperature 15 °C or lower.
• Xerophiles: growth at very low water activity (drought or near drought conditions).
• Thermophiles and hyperthermophiles: organisms that grow best at 40°C or higher, or 80°C or higher, respectively. Pyrolobus fumarii, a hyperthermophile, can live at a temperature 113°C. Another thermophile Thermus aquaticus, noted for its role in developing the polymerase chain reaction, is shown below.

• Toxicolerants: grow in the presence of high levels of damaging elements (e.g., pools of benzene, nuclear waste).
Archaea were originally seen as oddities of life, thriving in unfriendly environments. They also include organisms living in less extreme environments, including soils, marshes and even in the human colon. They are also abundant in the oceans where they are a major part of plankton, participating in the carbon and nitrogen cycles. In the guts of cows, humans and other mammals, methanogens facilitate digestion, generating methane gas in the process. In fact, cows have even been cited as a major cause of global warming because of their prodigious methane emissions! On the plus side, methanogenic Archaea are being exploited to create biogas and to treat sewage. Other extremophiles are the source of enzymes that function at high temperatures or in organic solvents. As already noted, some of these have become part of the biotechnology toolbox.
C. The Eukaryotes
1. Large Compartmentalized Cells
The volume of a typical eukaryotic cell is some 1000 times that of a typical bacterial cell. Eukaryotic life would not even have been possible if not for a division of labor of eukaryotic cells among different organelles (membrane-bound structures). Imagine a bacterium as a 100 square foot room (the size of a small bedroom, or a large walk-in closet!) with one door. Now imagine a room 1000 times as big. That is, imagine a 100,000 square foot ‘room’. You would expect many smaller rooms inside such a large space, each with its own door(s). The eukaryotic cell is a lot like that large space, with lots of interior “rooms” (i.e., organelles) with their own entryways and exits. The smaller prokaryotic “room” has a much larger plasma membrane surface area/volume ratio than a typical eukaryotic cell, enabling required environmental chemicals to enter and quickly diffuse throughout the cytoplasm of the bacterial cell. Chemical communication between parts of a small cell is therefore rapid. In contrast, the communication over a larger expanse of cytoplasm inside a eukaryotic cell requires the coordinated activities of subcellular compartments. Such communication might be relatively slow. In fact, eukaryotic cells have lower rates of metabolism, growth and reproduction than do prokaryotic cells. The existence of large cells required the evolution of a division of labor supported by compartmentalization.
2. Animal and Plant Cell Structure Overview
Typical animal and plant cells with their organelles and other structures are illustrated below.
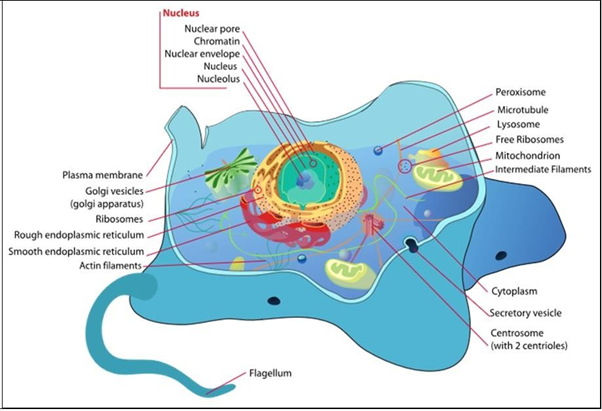
A plasma (cell) membrane surrounds all cells. A cell wall further surrounds prokaryotic, algal, fungal and plant cells, creating rigid structure around the cell membrane and supporting cell shape. Bacterial cell walls are composed of peptidoglycan, long polysaccharide chains attached to polypeptide (amino acid) chains. Cellulose, hemicellulose, and pectin are major polysaccharides of the plant cell wall. Fungal cells contain a wall, whose principal component is chitin. Chitin is the same material that makes up the exoskeleton or arthropods (including insects and lobsters!). Fungi, more closely related to animal than plant cells, are a curious beast for a number of reasons! For one thing, the organization of fungi and fungal cells is somewhat less defined than animal cells. Structures between cells called septa separate fungal hyphae, allow passage of cytoplasm and even organelles between cells. Some primitive fungi have few or no septa, in effect creating coenocytes, which are single giant cells, with multiple nuclei.
We end this look at the domains of life by noting that, while eukaryotes are a tiny minority of all living species, “their collective worldwide biomass is estimated at about equal to that of prokaryotes” (Wikipedia). On the other hand, our bodies contain 10 times as many microbial cells as human cells! In fact, it is becoming increasingly clear that a human owes as much of its existence to its microbiota (see above) as it does to its human cells. Keeping in mind that plants and animal cells share many internal structures and organelles that perform the same or similar functions, let’s look at them and briefly describe their functions.