
2.6.1: Introduction to Seed Plants
- Page ID
- 47643
Learning Objectives
- Explain how heterospory led to the evolution of seeds.
- Connect r- and k-strategies to propagules produced by plants (e.g. spores, gametes, pollen, and seeds).
- List a few ways gymnosperms and angiosperms differ in life history traits.
Fossil from the Devonian period reveal fernlike plants that were heterosporous; that is, produced two kinds of spores: microspores and megaspores. The megaspores were not released from the parent sporophyte. Fertilization took place within the tissue of the parent sporophyte thus freed from dependence on surface water. However, the necessity for the microspores to be carried from one plant to another in order to reach the megagametophyte robbed them of their value as agents of dispersal. This function was taken over by seeds - dormant, protected, embryo sporophytes.
Heterospory: The Next Big Step for Life on Land
Vertebrate animals became fully terrestrial only when their fertilization became completely independent from water. Plants started to perform the similar “evolutionary efforts” even earlier, but while reptiles actively approach the sexual partner, plants evolved different solutions. Instead of the active sex, plants use “carpet bombing” with spores; this increased the chance that two spores land nearby and the distance between sperm and egg cell will be minimal.
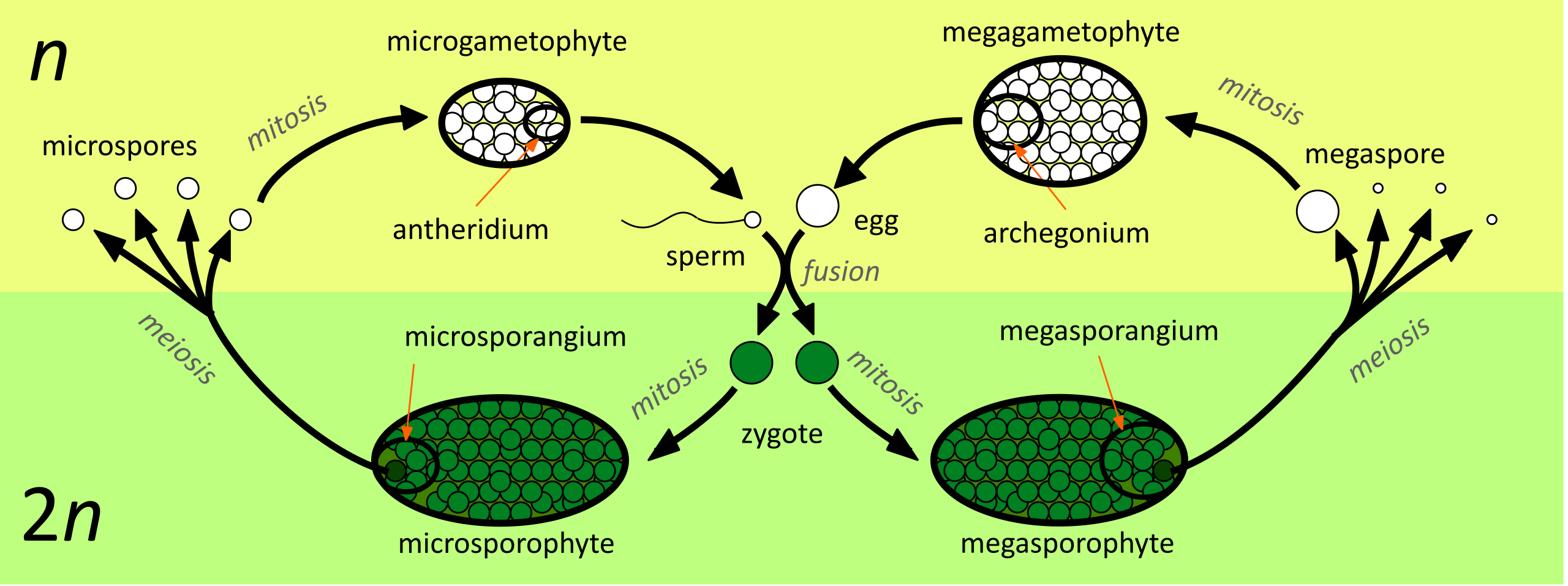
However, increasing the number of spores also results in a waste of resources, so plants minimized spore size; this will also allow for the longer distance of dispersal and fewer resources invested in each spore. On the other hand, some spores must remain large because the embryo (if fertilization occurs) will need the support from the feeding gametophyte. Consequently, plants ended up with division of labor: numerous, minuscule microspores that grow into microgametophytes with antheridia only, and a few large megaspores that make megagametophytes producing only archegonia (Figure \(\PageIndex{1}\)). Megaspores are larger because they are rich in nutrients, ready to be fertilized and nourish a developing sporophyte.

This heterosporic cycle makes fertilization less dependent on water and more dependent on spore distribution and gametophyte features (Figure \(\PageIndex{2}\)). It also allows for increased variations for selection to act upon. Division of labor allows resources to be used more efficiently and also restricts self-fertilization. In plant evolution, there was a high need for heterospory because it independently arose in several groups of seedless vascular plants (e.g. Selaginella and Azolla) and even among mosses.
As the evolutionary trajectory of heterospory progresses, some megaspores do not leave the mother plant and instead germinate there, waiting for the fertilization from a nearby microgametophyte: a step towards the first seeds. Male gametophytes become so small that they can easily be transported as a whole. Entire male gametophytes, rather than just gametes, start to be a mobile stage—this is origin of pollination.
Getting Larger: Secondary Growth and the Transition From Seeds to Spores
When plants developed secondary growth, a type of growth that allowed for lateral expansion and accumulation of woody tissue, they were able to grow much larger. However, these giants faced a new problem. Big animals like elephants, lions, and whales tend to produce a small number of offspring but increase the child care to ensure survival. Another strategy, usually employed by smaller organisms, is to produce a large number of offspring with low investment in each (such as most fungal spores), though most of them will not survive (Figure \(\PageIndex{3}\)).

Imagine these big, secondary thickening spore plants: they made billions of spores with little nutrients or protection for the developing offspring. Naturally, only few from these billions would survive to become fertilized. Spore reproduction is cheap and efficient but results are unpredictable. Even worse, these spore tree forests were not at all stable: in accidentally good conditions, many spores would survive and make sporophytes, which would all start to grow simultaneously and then suppress each other. But if the environmental conditions are poor, then none of the gametophytes will survive so there would be no new saplings to replace the old trees.
Plants evolved a solution to this conundrum: the seed. The idea of a seed is to hide most of the heterosporous life cycle inside the parent plant. In seed plants, everything happens directly on the parent sporophyte: development of gametophytes, syngamy, and growing of offspring sporophyte. Consequently, the megaspore never leaves the sporangium. It germinates inside, waits for fertilization and then the zygote grows into and embryo, still inside the same sporangium. What will finally leave the mother plant is the entire megasporangium with the megagametophyte and embryo inside. This is the seed's chimeric construction: sporophyte, megagametophyte, embryo, and (in angiosperms) endosperm genotypes encased within a single structure.
One problem is still left. How will sperm reach the retained megagametophyte and egg cell? The target is now perhaps high above the ground, on a branch of the giant tree. The only possible solution is pollination. Pollination is the distribution of the whole male gametophytes, also called pollen. Plants aren't able to physically go find a mate, so they always need a third party to transport their pollen, usually wind or insects.
Once the pollen gets to the intended destination, how would the sperm then swim to the egg cell? Some seed plants excrete a drop of liquid from the top of the ovule (a pollen drop) to create a fluid connection between the pollen grain to the ovule. Another sperm delivery tool, the pollen tube (Figure \(\PageIndex{4}\)), is made from one of the pollen grain cells in some plants. Fertilization with pollen tube is often called siphonogamy.
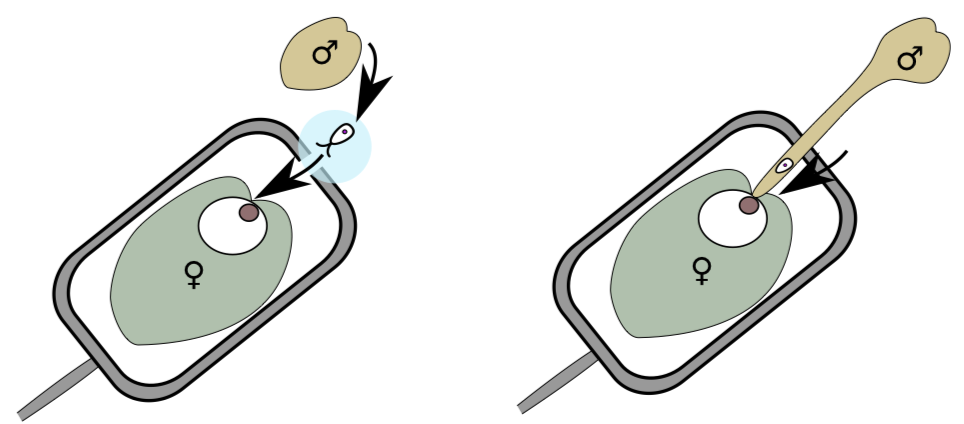
Consequently, seed plants with the pollen tube do not have flagella on male gametes; these cells are spermatia: non-motile male gametes. A pollen tube allows only two male gametes per gametophyte. Male gametes are usually competing for fertilization—this selects the best genotypes; whereas in higher seed plants, competition is between pollen tubes. The haploid pollen tube grows inside alien tissue of diploid sporophyte, so this growth is extremely slow in many seed plants. However, angiosperms made their pollen tubes grow fast.
With all these revolutionary adaptations, seed plants were first to colonize really dry places, and, in turn, allowed all other life to survive in arid climates. Read more about the evolution of seeds in this article from September, 2020.
Gymnosperms and Angiosperms
Seed plants are composed of two major lineages: gymnosperms (meaning naked seed) and angiosperms (meaning covered seed). These two lineages represent major differences in life history traits connected to the evolutionary conditions during which each evolved.
Gymnosperm reproduction differs from that of angiosperms in several ways. In gymnosperms, the megagametophyte is contained within an ovule, present on exposed bracts of the megastrobilus, where seeds develop and are often passively released (though some are dispersed by animals). In angiosperms, the ovule is contained within an enclosed ovary. The ovary then develops into a fruit, specialized for seed protection and dispersal. Double fertilization is a key event in the life cycle of angiosperms, forming a second supply of nutrients for the developing embryo (the endosperm), but is absent in most gymnosperms. The gametophyte structures are produced in separate strobili in gymnosperms, whereas in angiosperms, they are often a part of the same flower. Wind plays an important role in pollination in gymnosperms because pollen is blown by the wind to land on the female cones. Although many angiosperms are also wind-pollinated, animal pollination is more common, resulting in specialized pollination syndromes present in flowers.
Attributions
Curated and authored by Maria Morrow, CC BY-NC, using the following sources:
- 32.1 Reproductive Development and Structure from Biology 2e by OpenStax (licensed CC-BY). Access for free at openstax.org.
- 19.1.5 Diversity and Evolutionary Relationships of the Plants from Biology by John. W. Kimball (licensed CC-BY)
- 7.5 The Origin of Trees and Seeds from Introduction to Botany by Alexey Shipunov (public domain)