
5.4: The Stem
- Page ID
- 18007

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)The stem is an axial organ of shoot. It has functions of support, transportation, photosynthesis, and storage. Stem has radial structure, no root hairs and grows continuously.
Morphology of the Stem
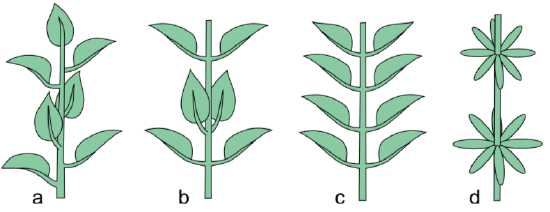
Stem morphology is simple. Its components are nodes (places where leaves are/were attached) and internodes, long or short (in the last case, plant sometimes appears to be stemless, rosette-like).Stems are different by the type of phyllotaxis. The phyllotaxis refers to the arrangement of leaves. If there is one leaf per node, it is a spiral (alternate) arrangement. Two leaves per node means opposite arrangement: two leaves per node arrangement. Opposite leaves can be all in the same plane or each pair can rotate at 90\(^\circ\). If there are more than two leaves per node, it is a whorled arrangement, and each whorl can also rotate. Each type of spiral phyllotaxis has its own angle of divergence. Multiple types of spiral leaf arrangement mostly follow the Fibonacci sequence:
\[\frac{1}{2}, \frac{1}{3}, \frac{2}{5}, \frac{3}{8}, \frac{5}{13}, \frac{8}{21}, ...\]
This sequence of numbers made with simple rule: in the every following fraction, the numerator and denominator are sums of two previous numerators and denominators, respectively. The sequence looks fairly theoretical but amazingly, it is fully applicable to plant science, namely to different types of spiral phyllotaxis (Figure \(\PageIndex{2}\)).
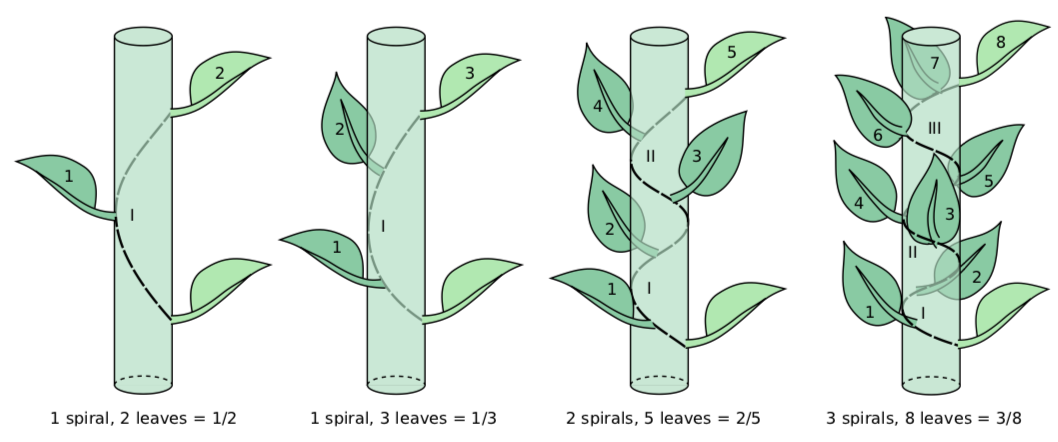
To determine formula of spiral phyllotaxis, one needs to start with arbitrary leaf (or leaf scar) and then find the next (upper) one which is directed the same way, lays on the same virtual line. Then, the imaginary spiral should be drawn trough basements from the started leaf to the corresponding upper leaf. This spiral should go through all intermediary leaves, there might be one, two or more intermediary leaves. Also, the spiral will go at least one time around the stem. (Instead of the imaginary spiral, it is sensible to use a thin thread). One needs to count all leaves in the spiral except the first, and also count number of rotations. The number of leaves counted will be the denominator of the formula, and the number of rotations is the numerator. This is how Fibonacci numbers appear in plant morphology.
These phyllotaxis formulas are relatively stable and sometimes even taxon-specific. For example, grasses (Gramineae) have \(\frac{1}{2}\) phyllotaxis, sedges (Carex) \(\frac{1}{3}\), many Rosaceae (like apple, Malus or cherry, Prunus) have \(\frac{2}{5}\), willows frequently have \(\frac{3}{8}\), et cetera. It is still not absolutely clear why the spiral phyllotaxis is under such a theoretical mathematical rule. The most feasible hypothesis emphasizes mathematical problem of circle packing and the competition between leaf primordia around SAM.
Anatomy of the Primary Stem
Plant evolution resulted first in the primary stems with no lateral meristems and secondary tissues. Only long after plants “learned” how to thicken their stems.
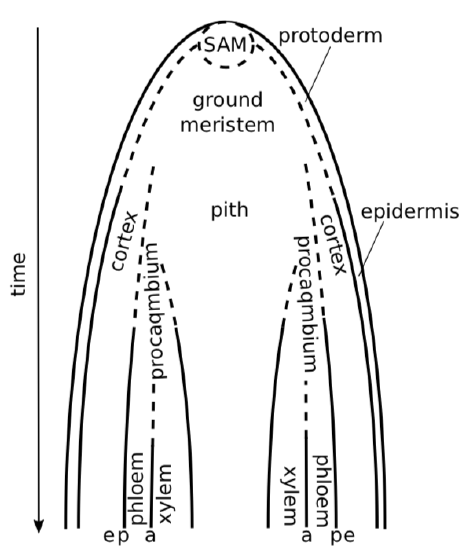
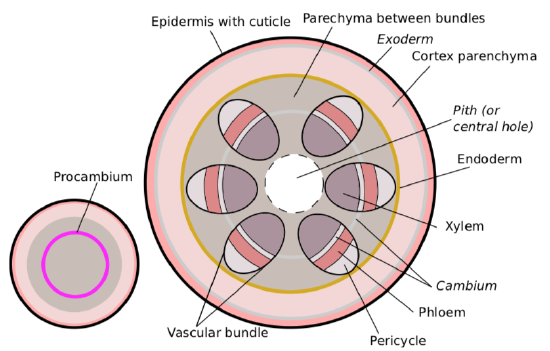
Development of stem starts from stem apical meristem (SAM) on the top of plant. The SAM produces three primary meristems: procambium, protoderm, and ground meristem. Protoderm cells differentiate into epidermal cells. The ground meristem differentiates into the cortexexternal layer of primary stem or root and pithcentral layer of primary stem or root. The procambium raises between the cortex and the pith. It forms vascular bundles or vascular cylinder.(Figure \(\PageIndex{2}\)).
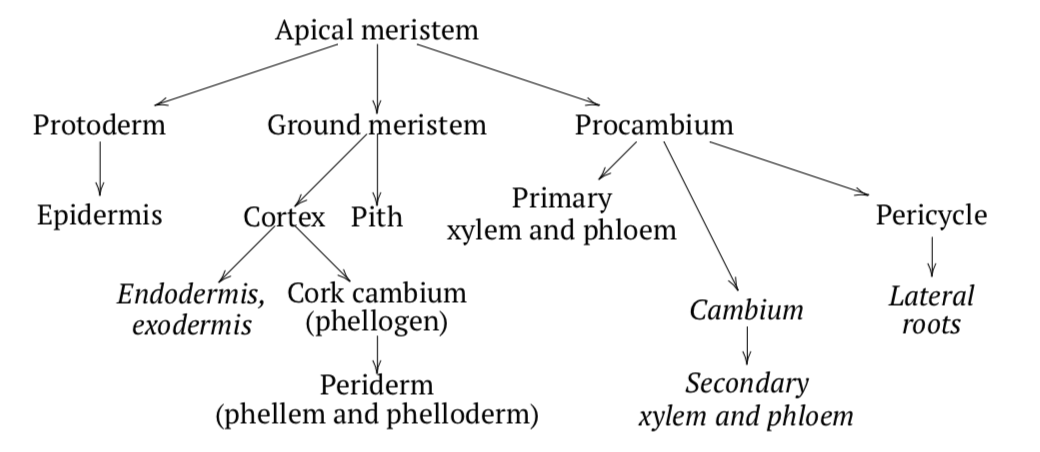
The outer layers of the procambium form the primary phloem. The inner layers become the primary xylem. The middle layer can be entirely spent or will make cambium for the secondary thickening. At times, the layers of the outside of the procambium can form a pericycle. Sometimes the innermost layer of the cortex can form an endodermis (endoderm) (Figure \(\PageIndex{3}\)), and outermost layer makes the exodermis (exoderm). All these layers are some kind of the “border control” between functionally different layers of stem. Another frequent variant is the development of collenchyma in the cortex adjacent to epidermis.
Vascular bundles connect leaves and stems. In many plants, they form a ring on the cross-section of the stem. Parenchyma (ground tissue) between vascular bundles typically belongs to both cortex and pith. Another variant is a vascular cylinder, structure which fully encircles the stem. Liliid (monocot) stems generally have dispersed vascular bundles. These three variants are steles, overall configurations of the primary vascular system of the plant stem (Figure 5.5.1). The most frequent kinds of steles are eustele (vascular bundles in a ring), solenostele (vascular cylinder) and ataktostele (dispersed vascular bundles).
All these types were probably originated from protostele, configuration where central xylem is surrounded with phloem and no pith is present (Figure 5.5.2). While the protostele was typical for many prehistoric plants, now only some lycophytes (Huperzia) have protostele in stems.
Saying that, it is important to note that roots of most plants have vascular tissues arranged similarly to protostele.