1.2: Introduction - Basic Chemistry
- Page ID
- 7802

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Source: BiochemFFA_1_2.pdf. The entire textbook is available for free from the authors at http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy
“Organic chemistry is the chemistry of carbon compounds. Biochemistry is the chemistry of carbon compounds that crawl” -Michael Adams.
To understand biochemistry, one must possess at least a basic understanding of organic and general chemistry. In this brief section, we will provide a rapid review of the simple concepts necessary to understand cellular chemistry. Chemistry is chemistry, whether in a cell or outside it, but biological chemistry is a particular subset of organic chemistry that often involves enormous macromolecules, and that happens in the aqueous environment of the cell.


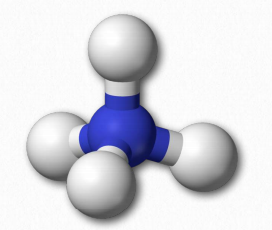
Covalent bonds, as you know, are the result of sharing of electrons between two atoms. Ionic bonds, by contrast, are formed when one atom donates an electron to another, such as in the formation of sodium chloride. Single covalent bonds can rotate freely, but double bonds cannot. Single bonds around a carbon atom are arranged in a tetrahedron with bond angles of 109.5° relative to each other, with the carbon at the center (Figure 1.19). Double bonded carbons create a planar structure with bond angles typically of about 120°.
Electronegativity
Electronegativity is a measure of the affinity a nucleus has for outer shell electrons (Table 1.2). High electronegativity corresponds to high affinity. Electrons in a covalent bond are held closer to the nucleus with a greater electronegativity compared to a nucleus with lower electronegativity.
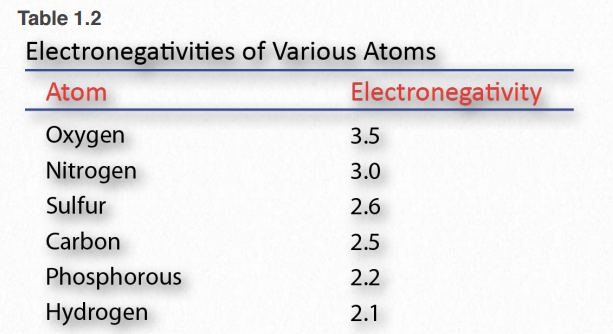
Table 1.2 Image by Aleia Kim
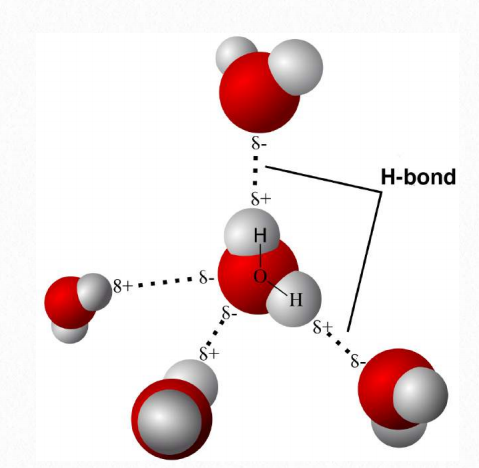
For example, in a molecule of water, with hydrogen covalently bonded to oxygen, the electrons are “pulled” toward the oxygen, which is more electronegative. Because of this, there is a slightly greater negative charge near the oxygen atom of water, compared to the hydrogen (which, correspondingly has a slightly higher positive charge). This unequal charge distribution sets up a dipole, with one side being somewhat negative and the other somewhat positive. Because of this, the molecule is described as polar.
Hydrogen bonds between water molecules are the result of the attraction of the partial positive and partial negative charges on different water molecules (Figure 1.20). Hydrogen bonds can also form between hydrogens with a partial positive charge and other strongly electronegative atoms, like nitrogen, with a partial negative charge. It is important to remember that hydrogen bonds are interactions between molecules (or parts of molecules) and are not bonds between atoms, like covalent or ionic bonds. Bonds between hydrogen and carbon do not form significant partial charges because the electronegativities of the two atoms are similar. Consequently, molecules containing many carbon-hydrogen bonds will not form hydrogen bonds and therefore, do not mix well with water. Such molecules are called hydrophobic. Other compounds with the ability to make hydrogen bonds are polar and can dissolve in water. They are called hydrophilic. Molecules possessing both characteristics are called amphiphilic.
Weak interactions
Hydrogen bonds are one kind of electrostatic (i.e., based on charge) interaction between dipoles. Other forms of electrostatic interactions that are important in biochemistry include weak interactions between a polar molecule and a transient dipole, or between two temporary dipoles. These temporary dipoles result from the movement of electrons in a molecule. As electrons move around, the place where they are, at a given time, becomes temporarily more negatively charged and could now attract a temporary positive charge on another molecule. Since electrons don’t stay put, these dipoles are very short-lived. Thus, the attraction that depends on these dipoles fluctuates and is very weak. Weak interactions like these are sometimes called van der Waals forces. Many molecular interactions in cells depend on weak interactions. Although the individual hydrogen bonds or other dipole-dipole interactions are weak, because of their large numbers, they can result in quite strong interactions between molecules.
Oxidation/reduction
Oxidation involves loss of electrons and reduction results in gain of electrons. For every biological oxidation, there is a corresponding reduction - one molecule loses electrons to another molecule. Oxidation reactions tend to release energy and are a source of bioenergy for chemotrophic cells.
Ionization
Ionization of biomolecules, by contrast does not involve oxidation/reduction. In ionization, a hydrogen ion (H+) leaves behind its electron as it exits (leaving behind a negative charge) or joins a group (adding a positive charge). Biological ionizations typically involve carboxyl groups or amines, though phosphates or sulfates can also be ionized. A carboxyl group can have two ionization states - a charge of -1 corresponds to the carboxyl without its proton and a charge of zero corresponds to the charge of the carboxyl with its proton on. An amine also has two ionization states. A charge of zero corresponds to a nitrogen with three covalent bonds (usually in the form of C-NH2) and a charge of +1 corresponds to a nitrogen making four covalent bonds (usually X-NH3 +).
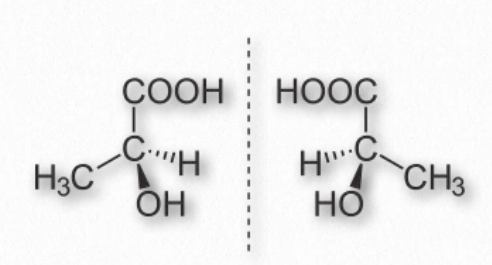
Stereochemistry
A carbon has the ability to make four single bonds (forming a tetrahedral structure) and if it bonds to four different chemical groups, their atoms can be arranged around the carbon in two different ways, giving rise to stereochemical “handedness” (Figure 1.21). Each carbon with such a property is referred to as an asymmetric center. The property of handedness only occurs when a carbon has four different groups bonded to it. Enzymes have very specific 3-D structures, so for biological molecules that can exist in different stereoisomeric forms, an enzyme that synthesizes it would make only one of the possible isomers. By contrast, the same molecules made chemically (not using enzymes) end up with equal amounts of both isomers, called a racemic mix.
Gibbs free energy
The Gibbs free energy calculation allows us to determine whether a reaction will be spontaneous, by taking into consideration two factors, change in enthalpy (ΔH) and change in entropy (ΔS). The free energy content of a system is given by the Gibbs free energy (\(G\)) and is equal to the enthalpy (\(H\)) for a process minus the absolute temperature (T) times the entropy (S)
\[G = H = TS\]
For a process, the change in the Gibbs free energy ΔG is given by
\[ΔG = ΔH - TΔS\]
A negative \(ΔG\) corresponds to release of free energy. Reactions that release energy are exergonic, whereas those that absorb energy are called endergonic.
The biological standard Gibbs free energy change (ΔG°’) corresponds to the ΔG for a process under standard conditions of temperature, pressure, and at pH = 7. For a reaction
\[aA + bB \rightleftharpoons cC + dD,\]
the equilibrium constant, \(K_{eq}\) is equal to
\[K_{eq} = \dfrac{ [C]^c_{eq} [D]^d_{eq}}{[A]^a_{eq} [B]^b_{eq}}\]
where \(a\), \(b\), \(c\), and \(d\) are integers in the balanced equation. Large values of \(K_{eq}\) correspond to favorable reactions (more C and D produced than A and B) and small values of \(K_{eq}\) mean the opposite. At equilibrium,
\[ΔG^{o\prime} = -RT \ln K_{eq}\]
If a process has a \(ΔG = Z\) and a second process has a \(ΔG = Y\), then if the two processes are linked, \(ΔG\) and \(ΔG^{o \prime} \) values for the overall reaction will be the sum of the individual ΔG and ΔG°’ values.
\[ΔG_{total} = ΔG_1+ ΔG_2 = Z + Y\]
\[ΔG^{o \prime }_{total} = ΔG_1^{o \prime}+ ΔG_2^{o\prime}\]
Catalysis
Catalysis is an increase in the rate of a reaction induced by a substance that is, itself, unchanged by the reaction. Because catalysts remain unchanged at the end of a reaction, a single catalyst molecule can be reused for many reaction cycles. Proteins that catalyze reactions in cells are called enzymes, while ribozymes are RNA molecules that act as catalysts.