3.1: Basic Concepts in Membranes
- Page ID
- 7815

Source: BiochemFFA_3_1.pdf. The entire textbook is available for free from the authors at http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy
Lipid bilayers
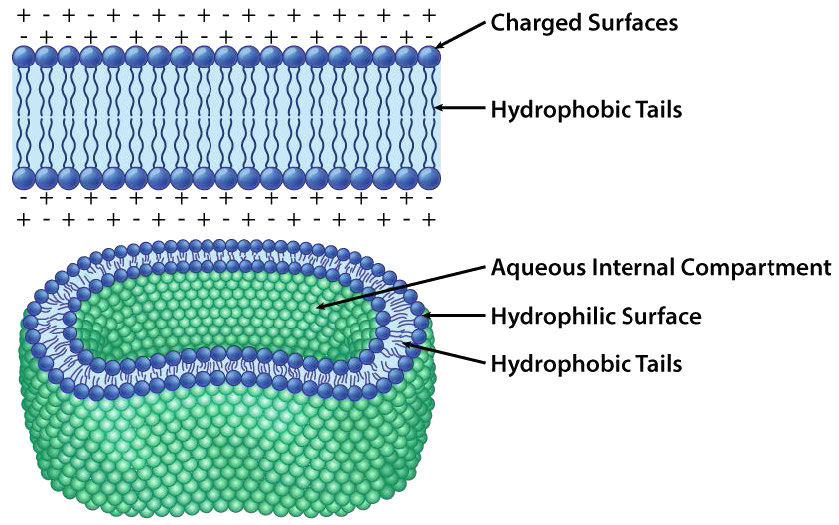
The protective membrane around cells contains many components, including cholesterol, proteins, glycolipids, glycerophospholipids, and sphingolipids. The last two of these will, when mixed vigorously with water, spontaneously form what is called a lipid bilayer (Figure 3.1), which serves as a protective boundary for the cell that is largely impermeable to the movement of most materials across it. With the notable exceptions of water, carbon dioxide, carbon monoxide, and oxygen, most polar/ionic require transport proteins to help them to efficiently navigate across the bilayer. The orderly movement of these compounds is critical for the cell to be able to 1) get food for energy; 2) export materials; 3) maintain osmotic balance; 4) create gradients for secondary transport; 5) provide electromotive force for nerve signaling; and 6) store energy in electrochemical gradients for ATP production (oxidative phosphorylation or photosynthesis). In some cases, energy is required to move the substances (active transport).
Facilitated Diffusion
In other cases, no external energy is required and they move by diffusion through specific cellular channels. This is referred to as facilitated diffusion. Before we discuss movement of materials across membranes, it is appropriate we discuss the composition of cellular membranes. Plasma membranes differ from cell walls both in the materials comprising them and in their flexibility. Cell walls will be covered near the end of this chapter.
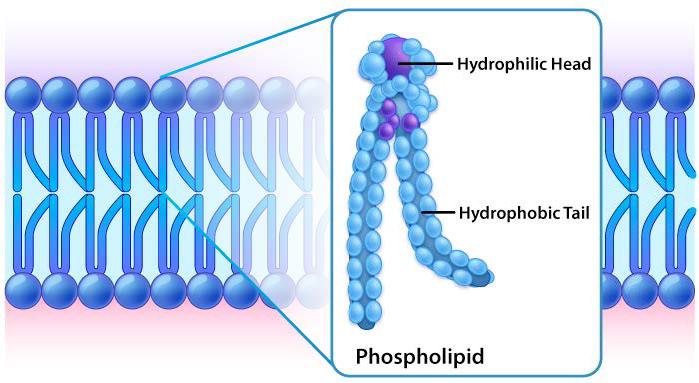
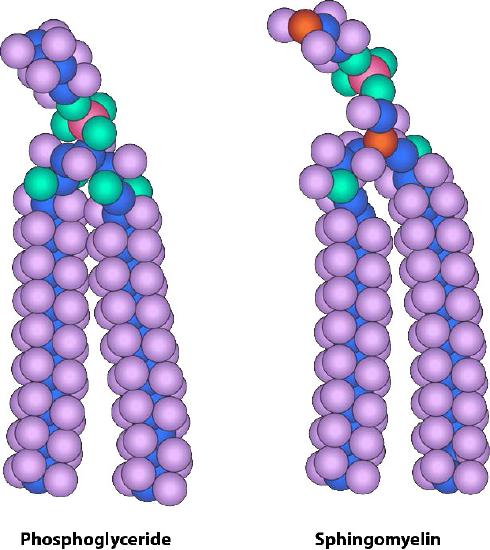
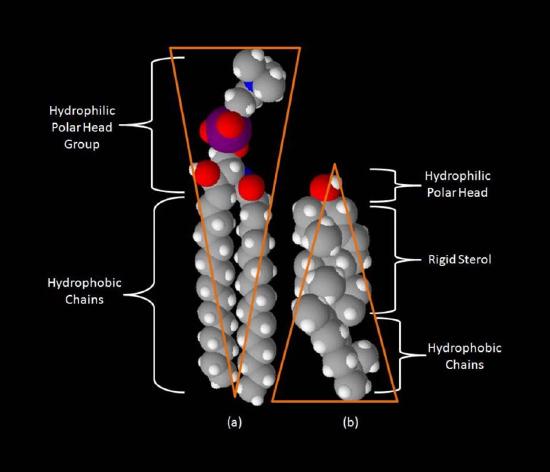
Though some cells do not have cell walls (animal cells) and others do (bacteria, fungi, and plants), there is commonality among cells in that they all possess plasma membranes. There is also commonality in the components of the membranes, though the relative amount of constituents varies. Figures 3.1 and 3.2 illustrate the structure and environments of plasma membranes. All plasma membranes contain a significant amount of amphiphilic substances linked to fatty acids. These include the glycerophospholipids and the sphingolipids. The fatty acid(s) are labeled as hydrophobic tails in the figures.
Hydrophilic heads
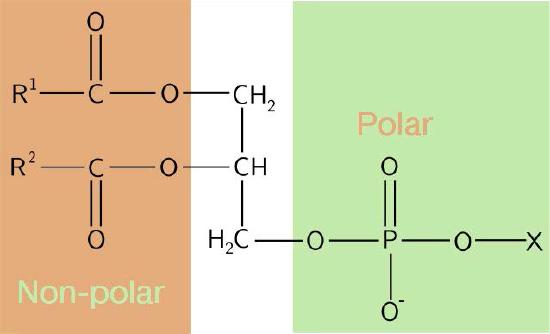
The composition of the hydrophilic heads varies considerably. In glycerophospholipids, a phosphate is always present, of course, and it is often esterified to another substance to make a phosphatide (Figure 3.3). Common compounds linked to the phosphate (at the X position) include serine, ethanolamine, and choline. These vary in the their charges so in this way, the charge on the external or internal surface can be controlled. Cells tend to have more negative charges on the exterior half of the lipid bilayer (called the outer leaflet) and more positive charges on the interior half (inner leaflet).
Sphingolipids
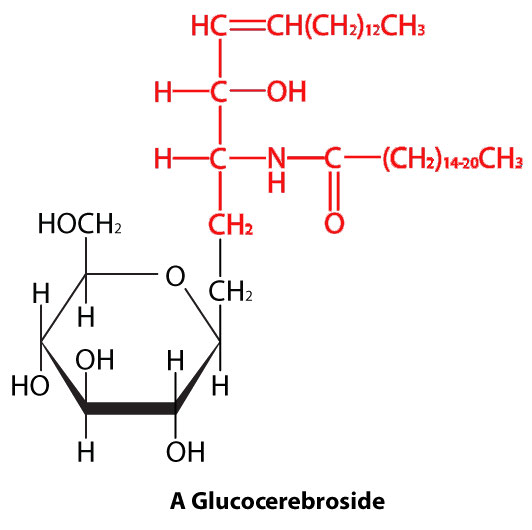
In sphingolipids (Figure 3.4), the hydrophilic head can contain a phosphate linked to ethanolamine or choline and this describes the structure of sphingomyelin, an important component of neural membranes. Most sphingolipids lack the phosphate and have instead a hydrophilic head of a single sugar (cerebrosides) or a complex oligosaccharide (gangliosides).
Water exclusion
In each case, the glycerophospholipid or sphingolipid has one end that is polar and one end that is non-polar. As we saw in the organization of amino acids with hydrophobic side chains occurring preferentially on the inside of a folded protein to exclude water, so too do the non-polar portions of these amphiphilic molecules arrange themselves so as to exclude water. Remember that the cytoplasm of a cell is mostly water and the exterior of the cell is usually bathed in an aqueous layer. It therefore makes perfect sense that the polar portions of the membrane molecules arrange themselves as they do - polar parts outside interacting with water and non-polar parts in the middle of the bilayer avoiding/excluding water.
Composition Bias
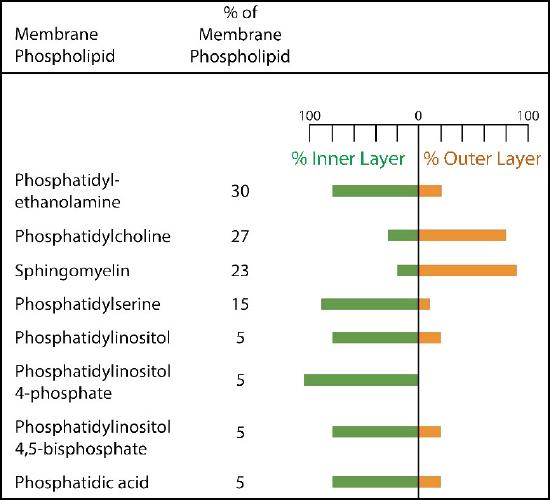
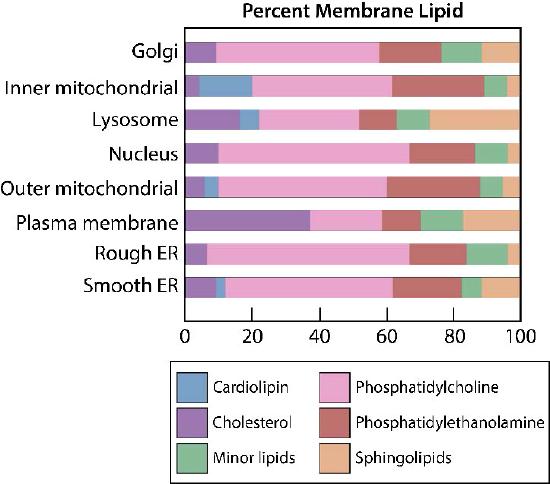
The plasma membrane has distinct biases of composition relative to its inside and the outside (Figure 3.7). First, glycosylation (of lipids and proteins) has the sugar groups located almost exclusively on the outside of the cell, away from the cytoplasm (Figure 3.8). Among the membrane lipids, sphingolipids are much more commonly glycosylated than glycerophospholipids. In addition, some of the glyerophospholipids are found preferentially on one side or the other (Figure 3.7). Phosphatidylserine and phosphatidylethanolamine are found preferentially within the inner leaflet of the plasma membrane, whereas phosphatidylcholine tends to be located on the outer leaflet. In the process of apoptosis, the phosphatidylserines appear on the outer leaflet where they serve as a signal to macrophages to bind and destroy the cell. Sphingolipids are found preferentially in the plasma membrane and are almost completely absent from mitochondrial and endoplasmic reticulum membranes (Figure 3.9).
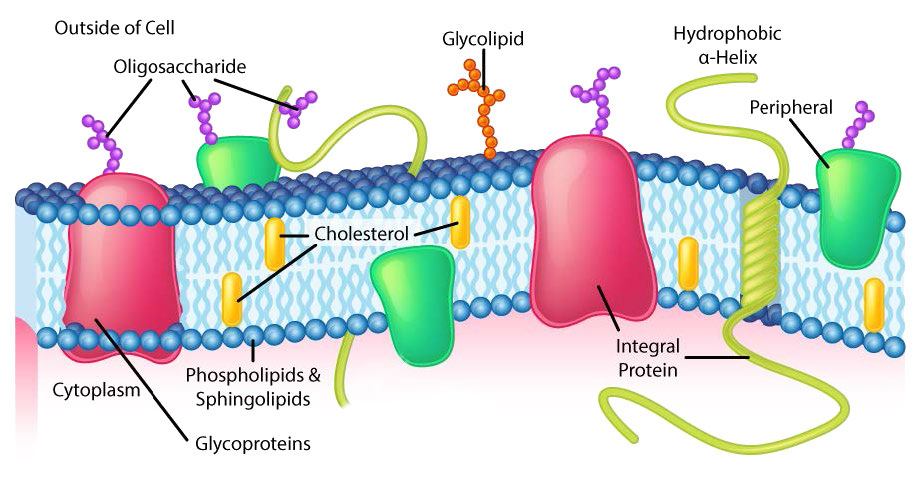
Organelle membranes
Bias of lipid composition also exists with respect to organelle membranes. The unusual diphosphodiglycerolipid known as cardiolipin, for example, is almost only found in mitochondrial membranes (see HERE) and like phosphatidylserine, its movement is an important step in apoptosis. In signaling, phosphatidylinositols play important roles providing second messengers upon being cleaved (see HERE).
Lateral Diffusion
Movement of lipids within each leaflet of the lipid bilayer occurs readily and rapidly due to membrane fluidity. This type of movement is called lateral diffusion and can be measured by the technique called FRAP (Figure 3.10, see HERE also). In this method, a laser strikes and stains a section of the lipid bilayer of a cell, leaving a spot as shown in B. Over time, the stain diffuses out ultimately across the entire lipid bilayer, much like a drop of ink will diffuse throughout when added to a glass of water. A measurement of the rate of diffusion gives an indication of the fluidity of a membrane.
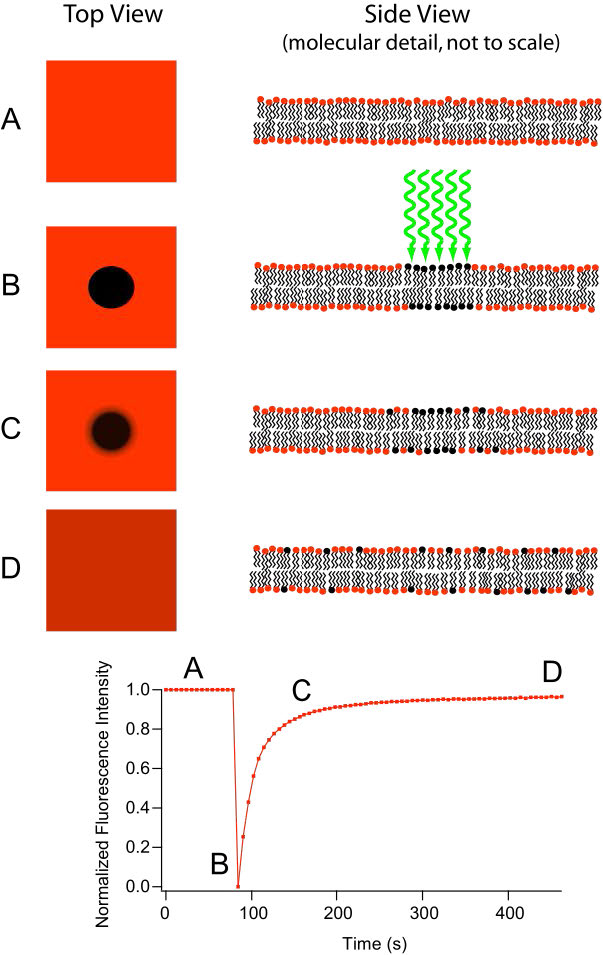
Transverse Diffusion
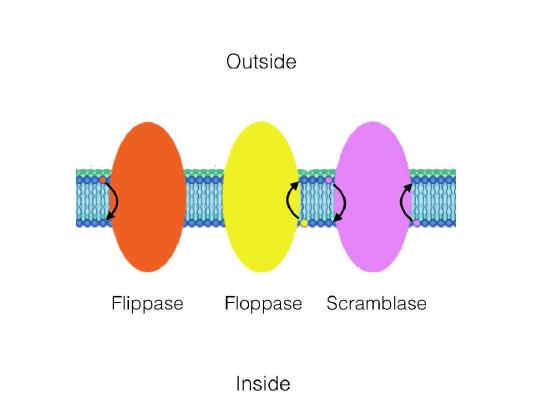
While the movement in lateral diffusion occurs rapidly, movement of molecules from one leaflet over to the other leaflet occurs much more slowly. This type of molecular movement is called transverse diffusion and is almost nonexistent in the absence of enzyme action. Remember that there is a bias of distribution of molecules between leaflets of the membrane, which means that something must be moving them.
There are three enzymes that catalyze movement of compounds in transverse diffusion. Flippases move membrane glycerophospholipids/ sphingolipids from outer leaflet to inner leaflet (cytoplasmic side) of cell. Floppases move membrane lipids in the opposite direction. Scramblases move in either direction.
Other components of lipid bilayer
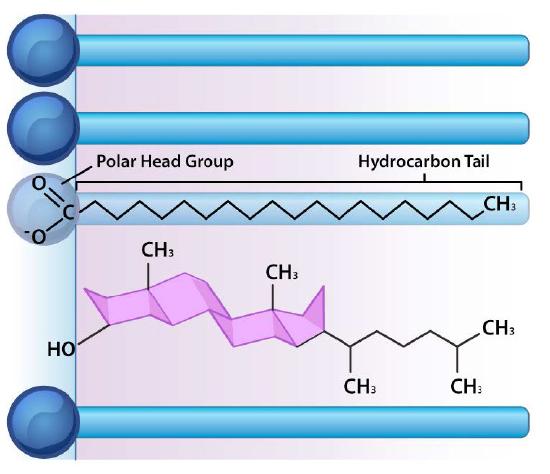
Besides glycerophospholipids and sphingolipids, there are other materials commonly found in lipid bilayers of cellular membranes. Two important prominent ones are cholesterol (Figure 3.13) and proteins. Besides serving as a metabolic precursor of steroid hormones and the bile acids, cholesterol’s main role in cells is in the membranes. The flatness and hydrophobicity of the sterol rings allow cholesterol to interact with the nonpolar portions of the lipid bilayer while the hydroxyl group on the end can interact with the hydrophilic part.
Membrane fluidity
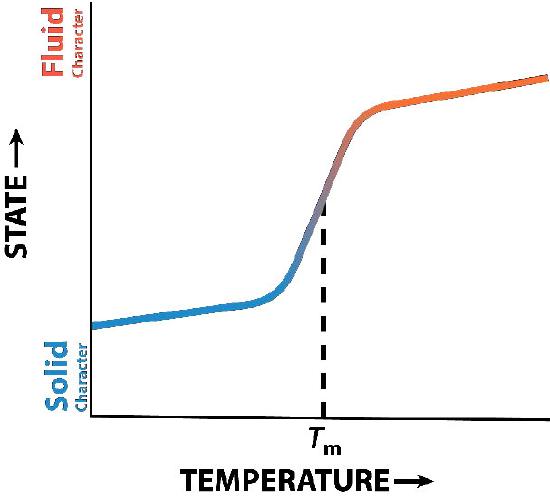
Cholesterol’s function in the lipid bilayer is complex (Figure 3.13). It influences membrane fluidity. Figure 3.14 shows the phase transition for a membrane as it is heated, moving from a more “frozen” character to that of a more “fluid” one as the temperature rises. The mid-point of this transition, referred to as the Tm, is influenced by the fatty acid composition of the lipid bilayer compounds. Longer and more saturated fatty acids will favor higher Tm values, whereas unsaturation and short fatty acids will favor lower Tm values. It is for this reason that fish, which live in cool environments, have a higher level of unsaturated fatty acids in them - to use to make membrane lipids that will remain fluid at ocean temperatures. Interestingly, cholesterol does not change the Tm value, but instead widens the transition range between frozen and fluid forms of the membrane, allowing it to have a wider range of fluidity.
Lipid Rafts
Cholesterol is also abundantly found in membrane structures called lipid rafts. Depicted in Figure 3.15, lipid rafts are organized structures within the membrane typically containing signaling molecules and other integral membrane proteins. Lipid rafts affect membrane fluidity, neurotransmission, and trafficking of receptors and membrane proteins.
Features
Distinguishing features of the rafts is that they are more ordered than the bilayers surrounding them, containing more saturated fatty acids (tighter packing and less disorganization, as a result) and up to 5 times as much cholesterol. They also are relatively rich in sphingolipids, with as much as 50% greater quantities of sphingomyelin than surrounding areas of the bilayer. The higher concentration of cholesterol in the rafts may be due to its greater ability to associate with sphingolipids (Figure 3.16). Some groups, such as prenylated proteins, like RAS, may be excluded from lipid rafts.
Lipid rafts may provide concentrating platforms after individual protein receptors bind to ligands in signaling. After receptor activation takes place at a lipid raft, the signaling complex would provide protection from nonraft enzymes that could inactivate the signal. For example, a common feature of signaling systems is phosphorylation, so lipid rafts might provide protection against dephosphorylation by enzymes called phosphatases. Lipid rafts appear to be involved in many signal transduction processes, such as T cell antigen receptor signaling, B cell antigen receptor signaling, EGF receptor signaling, immunoglobulin E signaling, insulin receptor signaling and others. For more on signaling, see HERE.
Barrier
Transport of materials across membranes is essential for a cell to exist. The lipid bilayer is an effective barrier to the entry of most molecules and without a means of allowing food molecules to enter a cell, it would die. The primary molecules that move freely across the lipid bilayer are small, uncharged ones, such as H2O, CO2, CO, and O2, so larger molecules, like glucose, that the cell needs for energy, would be effectively excluded if there were not proteins to facilitate its movement across the membrane.
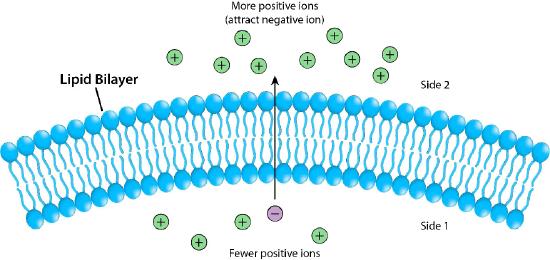
Figure 3.17 depicts the barrier that the lipid bilayer provides to movement across it and the pressures (ionic attraction, in this case) that can affect movement. Potential energy from charge and concentration differences are harvested by cells for purposes that include synthesis of ATP, and moving materials against a concentration gradient in a process called active transport.
Membrane proteins
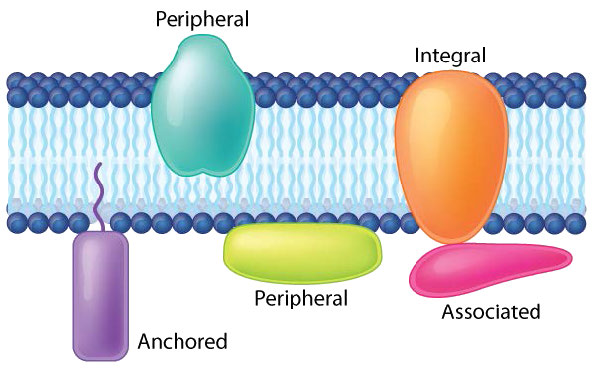
Proteins in a lipid bilayer can vary in quantity enormously, depending on the membrane. Protein content by weight of various membranes typically ranges between 30 and 75% by weight. Some mitochondrial membranes can have up to 90% protein. Proteins linked to and associated with membranes come in several types.
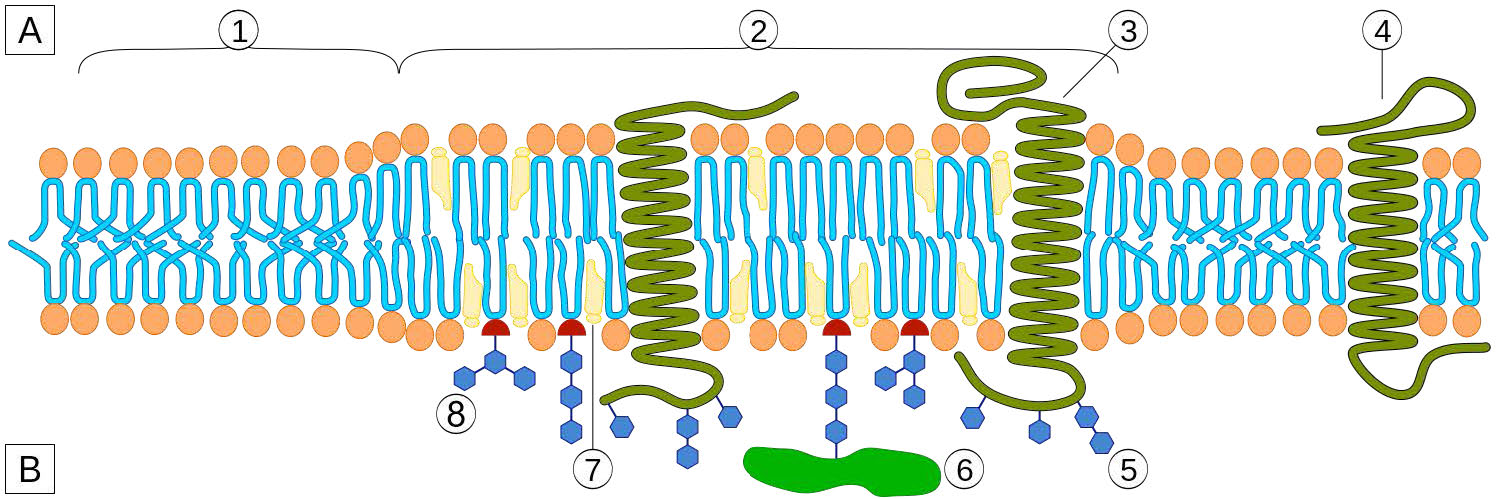
Transmembrane proteins
Transmembrane proteins are integral membrane proteins that completely span from one side of a biological membrane to the other and are firmly embedded in the membrane (Figure 3.18). Transmembrane proteins can function as docking sites for attachment (to the extracellular matrix, for example), as receptors in the cellular signaling system, or facilitate the specific transport of molecules into or out of the cell.
Example of integrated/ transmembrane proteins include those involved in transport (e.g., Na+/K+ ATPase), ion channels (e.g., potassium channel of nerve cells) and signal transduction across the lipid bilayer (e.g., GProtein Coupled Receptors).
Peripheral membrane proteins interact with part of the bilayer (usually does not involve hydrophobic interactions), but do not project through it. A good example is phospholipase A2, which cleaves fatty acids from glycerophospholipids in membranes. Associated membrane proteins typically do not have external hydrophobic regions, so they cannot embed in a portion of the lipid bilayer, but are found near them. Such association may arise as a result of interaction with other proteins or molecules in the lipid bilayer. A good example is ribonuclease.
Anchored membrane proteins
Anchored membrane proteins are not themselves embedded in the lipid bilayer, but instead are attached to a molecule (typically a fatty acid) that is embedded in the membrane (Figure 3.19). The oncogene family of proteins known as ras are good examples. These proteins are anchored to the lipid bilayer by attachment to non-polar farnesyl groups catalyzed by the enzyme farnesyltransferase.
Finer classification
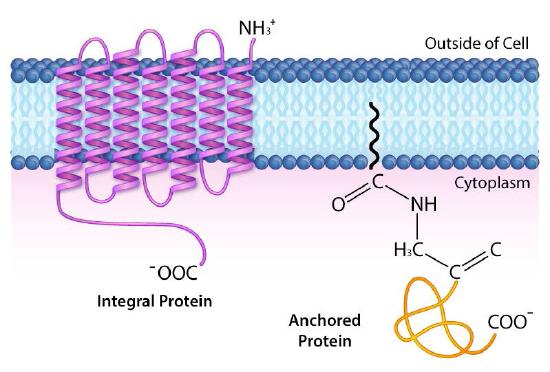
A more detailed classification scheme further categorizes the integral and anchored proteins into six different types (Figure 3.20). Type I and Type II have only one portion of the protein pass through the membrane. They differ in the orientation of the amine and carboxyl end with respect to inside/outside. Type I transmembrane proteins have the amino terminus on the outside and carboxy terminus on the inside, whereas Type II proteins have this reversed. Type III proteins are a single polypeptide chain that has multiple regions of it cross back and forth across the membrane, often to form a channel. Type IV is a multi-polypeptide protein which has multiple crossings of the membrane. Type V transmembrane proteins do not have a part of them that crosses the membrane, but they are anchored to the membrane by a lipid (such as a fatty acid) embedded in the lipid bilayer. Type VI transmembrane proteins both have a portion of them that crosses the membrane and they are attached to a lipid embedded in the lipid bilayer.
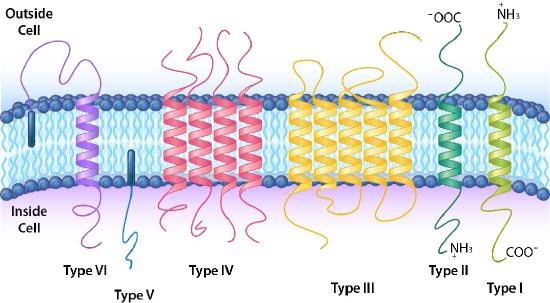
Blood Types
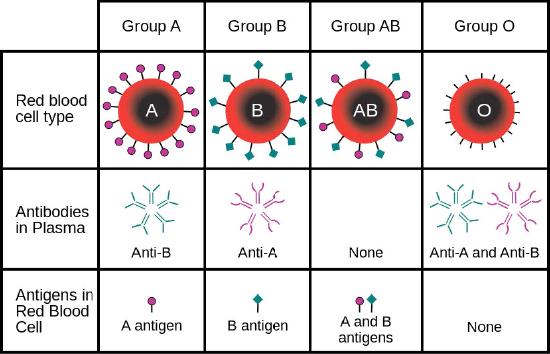
Cells have hundreds-thousands of membrane proteins and the protein composition of a membrane varies with its function and location. Glycoproteins embedded in membranes play important roles in cellular identification. Blood types, for example, differ from each other in the structure of the carbohydrate chains projecting out from the surface of the glycoprotein in their membranes (Figure 3.21).
Osmotic Pressure
Membranes provide barriers/boundaries for most molecules, but the permeability of water across a lipid bilayer creates a variable that must be considered. The variable here is osmotic pressure. Osmotic pressure (loosely) refers to the tendency of a solution to take in water by the process of osmosis. In Figure 3.22, one can see a visual representation of the concept of the pressure.
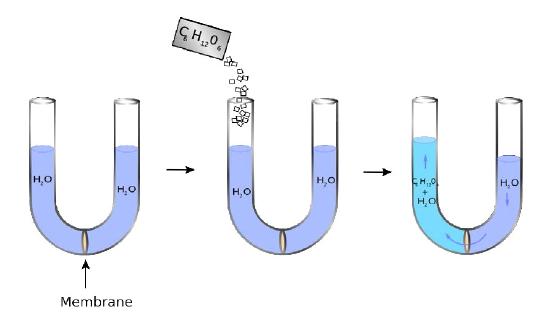
A U-shaped tube has at its bottom a semipermeable membrane. Water can pass through the membrane, but sugar molecules (C6H12O6) cannot. On the left side, sugar is added creating a concentration difference between the right and left chambers. Water diffuses across the membrane from right to left in an attempt to equalize the concentrations, causing the level of the right side to decrease and the left side to increase. The pressure resulting from the differences in height is felt at the membrane.
Equalizing concentrations
The liquid on the right does not completely move to the left, though, as might be expected if the only force involved is equalizing the concentration of sugar across the membrane (no sugar on right = no water). Instead, an equilibrium of sorts of water levels is reached even though the concentrations don’t equal out. The pressure existing at the membrane then from the differences in level corresponds to the osmotic pressure of the mixture. The osmotic pressure of a solution is the pressure difference needed to halt the flow of solvent across a semipermeable membrane. Osmotic pressure can also be thought of as the pressure required to counter osmosis. The osmotic pres- Figure 3.21 - Blood types arise from cell surface glycoproteins Figure 3.22 - Osmotic pressure. Water diffuses leftwards to try to equalize the solute concentration. The pressure realized at the membrane in the right figure is the osmotic pressure sure of a dilute solution mathematically behaves like the ideal gas law
\[P_{osmotic} = \dfrac{nRT}{V}\]
where n is the number of moles, R is the gas constant, T is the temperature in Kelvin, and V is the volume.
It is more convenient in solutions to work with molarity, so
\[P_{osmotic}= MR^* T\]
where M is the molarity of the dissolved molecules, R* is the gas constant expressed in (L atm)/(K mol), and T is the temperature. The Greek letter Π is used to refer to the Posmoticterm, so
\[Π = MR^* T\]
Remember when calculating the molarity to include the molarity of each particle. For example, when one dissolves sucrose in solution, it does not split into smaller particles, so
\[Molarity_{Particles} = Molarity_{Sucrose}\]
However, for salts, like KOH, which forms two ions in solution (K+ and OH- ),
MolarityParticles= 2* MolarityKOH.
Significant consideration
Osmotic pressure is a significant consideration for cells. Consider the fact that water can move freely across cellular membranes, but most of the contents of the cell, such as proteins, DNA, ions, sugars, etc., cannot. Second, the concentration of these compounds inside the cell is different than the concentration of them outside of the cell. Third, since water can move through the lipid bilayer, it will tend to move in the direction that will tend to equalize solute.
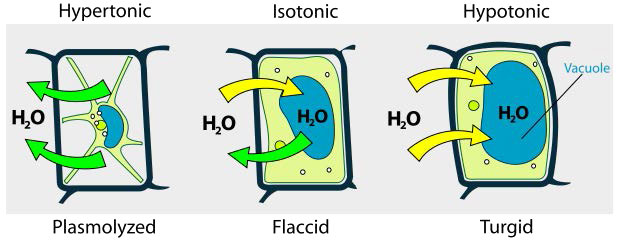
Hypotonic, hypertonic, isotonic
We consider three situations (Figure 3.23). First, if the concentration of solutes is greater inside the cell than outside, water will tend to move into the cell, causing the cell to swell. This circumstance is called hypotonic. Conversely, if the solute concentration is greater outside the cell than inside of it, water will exit the cell and the cell with shrink. This is a hypertonic situation. Last, if the concentrations of solutes into and outside of the cell is equal, this is called an isotonic solution. Here, no movement of water occurs across the cell membrane and the cell retains its size.
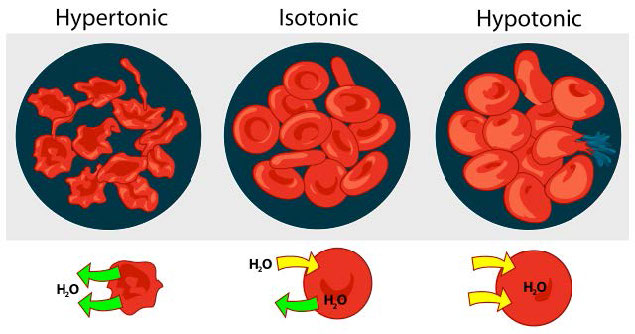
If the osmotic pressure is greater than the forces holding together a cellular membrane, the cell will rupture. Because of this, some cells have built in defenses to prevent problems. Plant cells, for example have a fairly rigid cell wall that resists expansion in hypotonic solutions (Figure 3.24). Bacteria also have a cell wall that provides protection.