
W2018_Bis2A_Lecture23_reading
- Page ID
- 25334
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Evolution and Natural Selection
Brief overview
Evolution and natural selection are core concepts in biology that are typically invoked to help explain the diversity of and relationships between life on Earth, both extant and extinct. Fortunately, in BIS2A, you need to understand and use only a few core ideas related to evolution and natural selection. We describe these below. You will expand your understanding and add details to these core concepts in BIS2B and BIS2C.
The first idea you need to grasp is that evolution can be simply defined as the development/change of something over time. In the automotive industry, the shapes and features of cars can be said to evolve (change in time). In fashion, it can be said that style evolves. In biology, life and, in particular, reproducing populations of organisms with different traits evolve.
The second thing to understand is that natural selection is a process by which nature filters organisms in a population. What is the filter? Here it becomes a little more complicated (but only a little). The simplest explanation is that the selective filter is just a combination of all living and nonliving factors in an environment, which influence how successfully an organism can reproduce. The factors that influence the ability of an organism to reproduce are known as selective pressures. A small but important complication is that these factors are not the same everywhere; they change in time and by location. Thus, the selective pressures that create the filter are constantly changing (sometimes rapidly, sometimes slowly), and organisms in the same reproducing population could experience different pressures at different times and in different locations.
The theory of evolution by natural selection puts these two ideas together; it stipulates that change in biology happens over time and that the variation in a population is constantly subjected to selection based on how differences in traits influence reproduction. But what are these characteristics or traits? What traits/features/functions can be subject to selection? The short answer is: just about anything associated with an organism for which variation exists in a population and for which this variation leads to a differential likelihood of generating offspring will probably be subject to filtering by natural selection. We also call these traits heritable phenotypes. Organisms in a population that have phenotypes, which enable them to pass the selective filter more efficiently than others, are said to have a selective advantage and/or greater fitness.
It is important to reiterate that while the phenotypes carried by individual organisms may be subject to selection, the process of evolution by natural selection both requires and acts on phenotypic variation within populations. If neither variation nor populations in which that variation can reside exist, there is no opportunity or need for selection. Everything is and stays the same.
Common misconceptions and a course specific note
Finally, we draw your attention to a critical point and common misconception among beginning students in biology. This misconception can arise when, for the sake of discussion, we decide to anthropomorphize nature by giving it an intellect. For example, we may try to build an example for evolution by natural selection by proposing that a surplus of a particular food exists in an environment and there is an organism close by that is starving. It would be correct to reason that if the organism could eat that food that this might give it a selective advantage over other organisms that cannot. If later we find an example of organisms that have the capability to eat that surplus food, it might be tempting to say that nature evolved to solve the problem the surplus food. The process of evolution by natural selection, however, happens randomly and without direction. That is, nature does NOT identify “problems” that are limiting fitness. Nature does NOT identify features that would make an organism more successful and then start creating diverse solutions that meet this need. The generation of variation is not guided. Variation happens and natural selection filters what works best. The observation that an organism exists that can eat the surplus food is not a reflection of nature actively solving a problem, but rather, a reflection of whatever processes that led to phenotypic variation in an ancestral population that created—among many other variants—a phenotype that increased fitness (possibly because the ancestral organisms were able to eat the surplus food).
This point of the preceding paragraph is particularly important to understand in the context of BIS2A because of the way we will be utilizing the Design Challenge to understand biology. While the Design Challenge is intended to help focus our attention on functions under selection and their relationship to determining fitness, it can be easy—if we aren’t attentive—to lapse into language that would suggest that nature purposefully designs solutions to solve specific problems. Always remember that we are looking retrospectively at what nature has selected and that we are attempting to understand why a specific phenotype may have been selected over many other possibilities. In doing so, we will be inferring or hypothesizing to the best of our ability (which is sometimes wrong) a sensible reason to explain why a phenotype might have provided a selective advantage. We are NOT saying that the phenotype evolved TO provide a specific selective advantage. The distinction between these two ideas may be subtle, but it is critical!
Note: possible discussion
What physical traits can you think of that give a selective advantage to certain species? Under what conditions would this trait grant those advantages? Under what conditions might that trait be a selective disadvantage?
Note: possible discussion
The great varieties of domesticated dog breeds from which we can choose for companionship are also the result of a process of evolution by selection. Likewise, the development of many very different looking crops—cabbage, brussel sprouts, kohlrabi, kale, broccoli and cauliflower—is also the result of evolution by selection. However, in these two cases the selection or filtering process is referred to artificial selection rather than natural selection. Discuss how artificial and natural selection are similar and different?
Note: possible discussion
How do environmental and political factors influence manufacturing processes such as automobile design? Fashion? Etc. What aspects are similar to the evolution of an organism, and what aspects are different?
Note: possible discussion
A related but slightly different misconception about evolution by natural selection is that this process leads to the creation of the most efficient solutions to problems. What is the problem with this notion?
Mutations
Errors occurring during DNA replication are not the only way by which mutations can arise in DNA. Mutations, variations in the nucleotide sequence of a genome, can also occur because of physical damage to DNA. Such mutations may be of two types: induced or spontaneous. Induced mutations are those that result from an exposure to chemicals, UV rays, x-rays, or some other environmental agent. Spontaneous mutations occur without any exposure to any environmental agent; they are a result of spontaneous biochemical reactions taking place within the cell.
Mutations may have a wide range of effects. Some mutations are not expressed; these are known as silent mutations. Point mutations are those mutations that affect a single base pair. The most common nucleotide mutations are substitutions, in which one base is replaced by another. These can be of two types, either transitions or transversions. Transition substitution refers to a purine or pyrimidine being replaced by a base of the same kind; for example, a purine such as adenine may be replaced by the purine guanine. Transversion substitution refers to a purine being replaced by a pyrimidine, or vice versa; for example, cytosine, a pyrimidine, is replaced by adenine, a purine. Mutations can also be the result of the addition of a nucleotide, known as an insertion, or the removal of a base, also known as deletion. Sometimes a piece of DNA from one chromosome may get translocated to another chromosome or to another region of the same chromosome; this is known as translocation.
As we will visit later, when a mutation occurs in a protein coding region it may have several effects. Transition or transversion mutants may lead to no change in the protein sequence (known as silent mutations), change the amino acid sequence (known as missense mutations), or create what is known as a stop codon (known as a nonsense mutation). Insertions and deletions in protein coding sequences lead to what are known as frameshift mutations. Missense mutations that lead to conservative changes results in the substitution of similar but not identical amino acids. For example, the acidic amino acid glutamate being substituted for the acidic amino acid aspartate would be considered conservative. In general we do not expect these types of missense mutations to be as severe as a non-conservative amino acid change; such as a glutamate substituted for a valine. Drawing from our understanding of functional group chemistry we can correctly infer that this type of substitution may lead to severe functional consequences, depending upon location of the mutation.
Note: Vocabulary Watch
Note that the preceding paragraph had a lot of potentially new vocabulary - it would be a good idea to learn these terms.
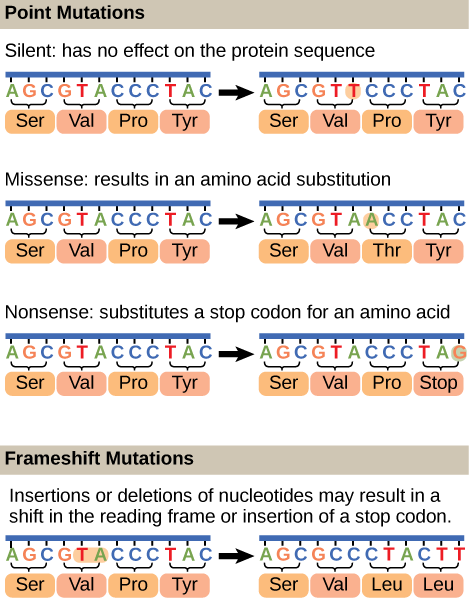
Figure 1. Mutations can lead to changes in the protein sequence encoded by the DNA.
Suggested discussion
Based on your understanding of protein structure, which regions of a protein would you think are more sensitive to substitutions, even conserved amino acid substitutions? Why?
Suggested discussion
A insertion mutation that results in the insertion of three nucleotides is often less deleterious than a mutation that results in the insertion of one nucleotide. Why?
Mutations: Some nomenclature and considerations
Mutation
Etymologically speaking, the term mutation simply means a change or alteration. In genetics, a mutation is a change in the genetic material - DNA sequence - of an organism. By extension, a mutant is the organism in which a mutation has occurred. But what is the change compared to? The answer to this question, is that it depends. The comparison can be made with the direct progenitor (cell or organism) or to patterns seen in a population of the organism in question. It mostly depends on the specific context of the discussion. Since genetic studies often look at a population (or key subpopulations) of individuals we begin by describing the term "wild-type".
Wild Type vs Mutant
What do we mean by "wild type"? Since the definition can depend on context, this concept is not entirely straightforward. Here are a few examples of definitions you may run into:
Possible meanings of "wild-type"
- An organism having an appearance that is characteristic of the species in a natural breeding population (i.e. a cheetah's spots and tear-like dark streaks that extend from the eyes to the mouth).
- The form or forms of a gene most commonly occurring in nature in a given species.
- A phenotype, genotype, or gene that predominates in a natural population of organisms or strain of organisms in contrast to that of natural or laboratory mutant forms.
- The normal, as opposed to the mutant, gene or allele.
The common thread to all of the definitions listed above is based on the "norm" for a set of characteristics with respect to a specific trait compared to the overall population. In the "Pre-DNA sequencing Age" species were classified based on common phenotypes (what they looked like, where they lived, how they behaved, etc.). A "norm" was established for the species in question. For example, Crows display a common set of characteristics, they are large, black birds that live in specific regions, eat certain types of food and behave in a certain characteristic way. If we see one, we know its a crow based on these characteristics. If we saw one with a white head, we would think that either it is a different bird (not a crow) or a mutant, a crow that has some alteration from the norm or wild type.
In this class we take what is common about those varying definitions and adopt the idea that "wild type" is simply a reference standard against which we can compare members of a population.
Suggested discussion
If you were assigning wild type traits to describe a dog, what would they be? What is the difference between a mutant trait and variation of a trait in a population of dogs? Is there a wild type for a dog that we could use as a standard? How would we begin to think about this concept with respect to dogs?

Mutations are simply changes from the "wild type", reference or parental sequence for an organism. While the term "mutation" has colloquially negative connotations we must remember that change is neither inherently "bad". Indeed, mutations (changes in sequences) should not primarily be thought of as "bad" or "good", but rather simply as changes and a source of genetic and phenotypic diversity on which evolution by natural selection can occur. Natural selection ultimately determines the long-term fate of mutations. If the mutation confers a selective advantage to the organism, the mutation will be selected and may eventually become very common in the population. Conversely, if the mutation is deleterious, natural selection will ensure that the mutation will be lost from the population. If the mutation is neutral, that is it neither provides a selective advantage or disadvantage, then it may persist in the population. Different forms of a gene, including those associated with "wild type" and respective mutants, in a population are termed alleles.
Consequences of Mutations
For an individual, the consequence of mutations may mean little or it may mean life or death. Some deleterious mutations are null or knock-out mutations which result in a loss of function of the gene product. These mutations can arise by a deletion of the either the entire gene, a portion of the gene, or by a point mutation in a critical region of the gene that renders the gene product non-functional. These types of mutations are also referred to as loss-of-function mutations. Alternatively, mutations may lead to a modification of an existing function (i.e. the mutation may change the catalytic efficiency of an enzyme, a change in substrate specificity, or a change in structure). In rare cases a mutation may create a new or enhanced function for a gene product; this is often referred to as a gain-of-function mutation. Lastly, mutations may occur in non-coding regions of DNA. These mutations can have a variety of outcomes including altered regulation of gene expression, changes in replication rates or structural properties of DNA and other non-protein associated factors.
Suggested discussion
In the discussion above what types of scenarios would allow such a gain-of-function mutant the ability to out compete a wild type individual within the population? How do you think mutations relate to evolution?
Mutations and cancer
Mutations can affect either somatic cells or germ cells. Sometimes mutations occur in DNA repair genes, in effect compromising the cell's ability to fix other mutations that may arise. If, as a result of mutations in DNA repair genes, many mutations accumulate in a somatic cell, they may lead to problems such as the uncontrolled cell division observed in cancer. Cancers, including forms of pancreatic cancer, colon cancer, and colorectal cancer have been associated with mutations like these in DNA repair genes. If, by contrast, a mutation in DNA repair occurs in germ cells (sex cells), the mutation will be passed on to the next generation, as in the case of diseases like hemophilia and xeroderma pigmentosa. In the case of xeroderma pigmentoas individuals with compromised DNA repair processes become very sensitive to UV radiation. In severe cases these individuals may get severe sun burns with just minutes of exposure to the sun. Nearly half of all children with this condition develop their first skin cancers by age 10.
Consequences of errors in replication, transcription and translation
Something key to think about:
Cells have evolved a variety of ways to make sure DNA errors are both detected and corrected, rom proof reading by the various DNA-dependent DNA polymerases, to more complex repair systems. Why did so many different mechanisms evolve to repair errors in DNA? By contrast, similar proof-reading mechanisms did NOT evolve for errors in transcription or translation. Why might this be? What would be the consequences of an error in transcription? Would such an error effect the offspring? Would it be lethal to the cell? What about translation? Ask the same questions about the process of translation. What would happen if the wrong amino acid was accidentally put into the growing polypeptide during the translation of a protein? Contrast this with DNA replication.
Mutations as instruments of change
Mutations are how populations can adapt to changing environmental pressures
Mutations are randomly created in the genome of every organism, and this in turn creates genetic diversity and a plethora of different alleles per gene per organism in every population on the planet. If mutations did not occur, and chromosomes were replicated and transmitted with 100% fidelity, how would cells and organisms adapt? Whether mutations are retained by evolution in a population depends largely on whether the mutation provides selective advantage, poses some selective cost or is at the very least, not harmful. Indeed, mutations that appear neutral may persist in the population for many generations and only be meaningful when a population is challenged with a new environmental challenge. At this point the apparently previously neutral mutations may provide a selective advantage.
Example: Antibiotic resistance
The bacterium E. coli is sensitive to an antibiotic called streptomycin, which inhibits protein synthesis by binding to the ribosome. The ribosomal protein L12 can be mutated such that streptomycin no longer binds to the ribosome and inhibits protein synthesis. Wild type and L12 mutants grow equally well and the mutation appears to be neutral in the absence of the antibiotic. In the presence of the antibiotic wild type cells die and L12 mutants survive. This example shows how genetic diversity is important for the population to survive. If mutations did not randomly occur, when the population is challenged by an environmental event, such as the exposure to streptomycin, the entire population would die. For most populations this becomes a numbers game. If the mutation rate is 10-6 then a population of 107 cells would have 10 mutants; a population of 108 would have 100 mutants, etc.
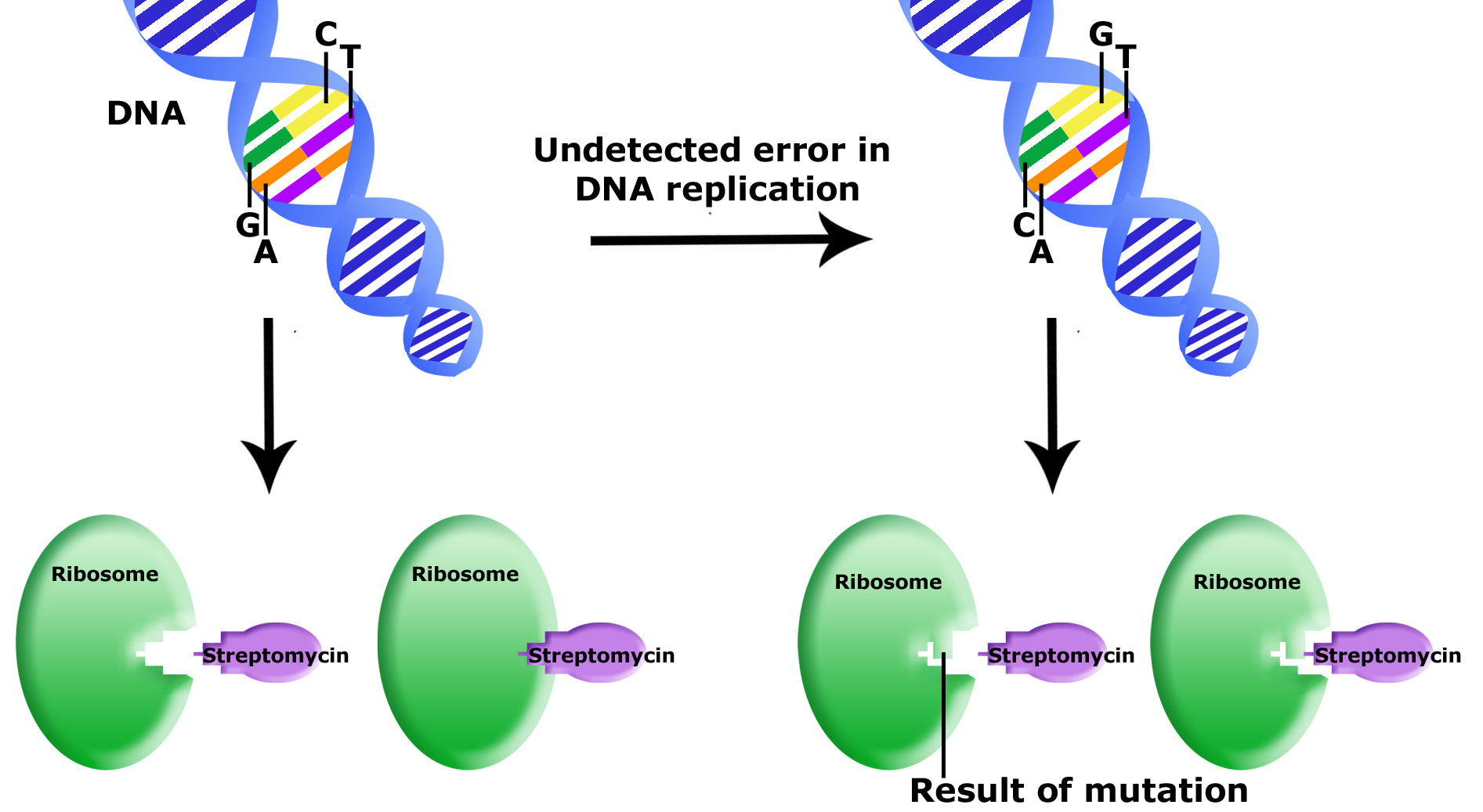
Uncorrected errors in DNA replication lead to mutation. In this example, an uncorrected error was passed onto a bacterial daughter cell. This error is in a gene that encodes for part of the ribosome. The mutation results in a different final 3D structure of the ribosome protein. While the wildtype ribosome can bind to streptomycin (an antibiotic that will kill the bacterial cell by inhibiting the ribosome function) the mutant ribosome cannot bind to streptomycin. This bacteria is now resistant to streptomycin.
Source: Bis2A Team original image
Suggested discussion
Based on our example, if you were to grow up a culture of E. coli to population density of 109 cells/ml; would you expect the entire population to be identical? How many mutants would you expect to see in 1 ml of culture?
An example: Lactate dehydrogenase
Lactate Dehydrogenase (LDH), the enzyme that catalyzes the reduction of pyruvate into lactic acid in fermentation, while virtually every organism has this activity, the corresponding enzyme and therefore gene differs immensely between humans and bacteria. The proteins are clearly related, they perform the same basic function but have a variety of differences, from substrate binding affinities and reaction rates to optimal salt and pH requirements. Each of these attributes have been evolutionarily tuned for each specific organism through multiple rounds of mutation and selection.
Suggested discussion
We can use comparative DNA sequence analysis to generate hypotheses about the evolutionary relationships between three or more organisms. One way to accomplish this is to compare the DNA or protein sequences of proteins found in each of the organisms we wish to compare. Let us, for example, imagine that we were to compare the sequences of LDH from three different organisms, Organism A, Organism B and Organism C. If we compare the LDH protein sequence from Organism A to that from Organism B we find a single amino acid difference. If we now look at Organism C, we find 2 amino acid differences between its LDH protein and the one in Organism A and one amino acid difference when the enzyme from Organism C is compared to the one in Organism B. Both organisms B and C share a common change compared to organism A.

Schematic depicting the primary structures of LDH proteins from Organism A, Organism B, and Organism C. The letters in the center of the proteins line diagram represent amino acids at a unique position and the proposed differences in each sequence. The N and C termini are also noted H2N and COOH, respectively.
Attribution: Marc T. Facciotti (original work)
Question: Is Organism C more closely related to Organism A or B? The simplest explanation is that Organism A is the earliest form, a mutation occurred giving rise to Organism B. Over time a second mutation arose in the B lineage to give rise to the enzyme found in Organism C. This is the simplest explanation, however we can not rule out other possibilities. Can you think of other ways the different forms of the LDH enzyme arose these three organisms?
Sickle cell anemia: a look at the connection between DNA and phenotype
Genes are translated into proteins; mutations often (but not always) result in changes in the sequence of amino acids in those proteins. Changes in the amino acid sequence can modify (in various ways) or even completely destroy protein function. Proteins have many functions within cells, and a change in those functions results in a change in the phenotype of that cell or organism. So a mutation as simple as a single base change in a DNA sequence can have dramatic effects on phenotype. One of the best examples of this phenomenon can be observed when mutations occur in the gene for one of the protein components of the red blood cell protein we call hemoglobin.
A major component of the erythrocytes (red blood cells) found in vertebrates is hemoglobin. A molecule of hemoglobin from a normal adult human contains four proteins (two identical alpha polypeptides and two identical beta polypeptides) surrounding a core of heme (complex molecule containing an atom of iron which can combine reversibly with oxygen). Thus, hemoglobin functions as the major oxygen-carrying constituent of blood. Because of hemoglobin, a given volume of blood can carry far more oxygen than could be dissolved in an equal volume of water.
In many human populations, particularly those with origins in Central Africa or the Mediterranean, there are individuals who suffer from severe anemia and whose blood contains numerous distorted, sickle-shaped erythrocytes. Hence, the disease was given the name sickle cell anemia.
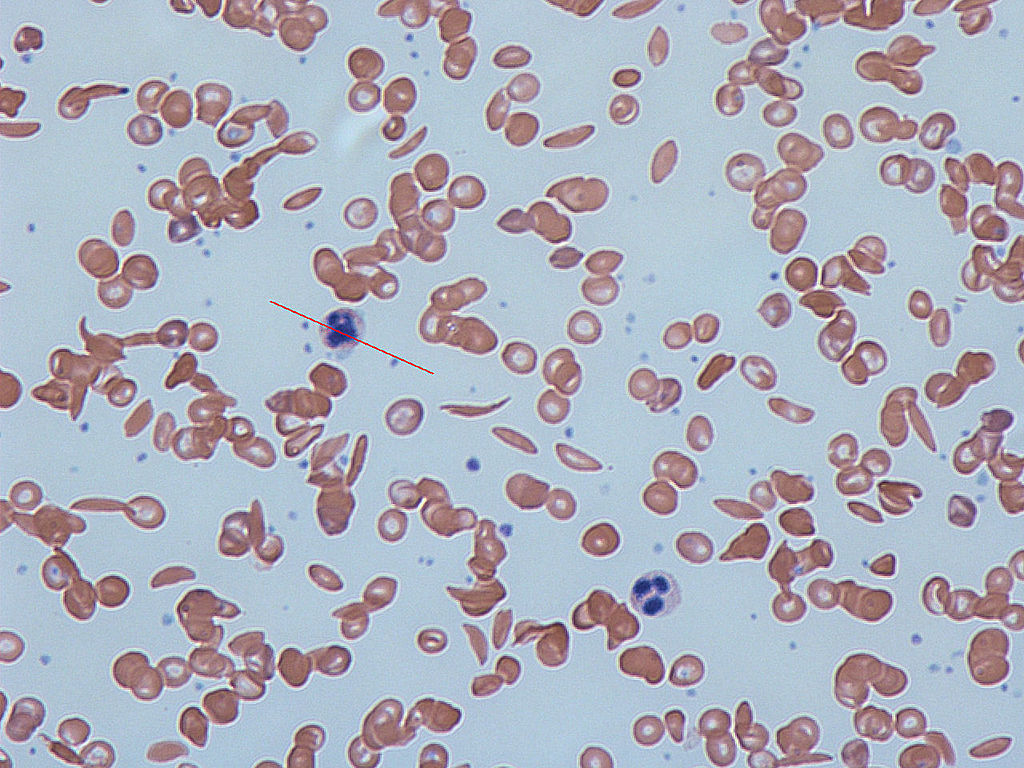
Figure 1. Notice the sickle shaped cells in the image by Dr Graham Beards via Wikimedia Commons
Biochemical studies established that the gene affected in sickle-cell anemia has the code for an abnormal beta polypeptide, which is one of the components of the hemoglobin molecule. Therefore, there are two different forms of the hemoglobin gene that codes for the beta chain:
- Form 1: HbA has the code for a normal beta chain
- Form 2: HbS has the code for an abnormal beta chain
Humans are diploid organisms; they have two copies of most genes. However, the two copies they possess do not have to be identical. When there are two possible alleles for a gene (such as in the gene for the beta chain of hemoglobin), a diploid individual will have one of three possible combinations of the two alleles. They can be HbA HbA , HbA HbS, or HbSHbS.
The set of alleles present in an individual for a given gene is known as the individual’s genotype. The three combinations of two alleles above are therefore the three different genotypes. Individuals that have two copies of the same allele are called homozygous; individuals with two different alleles are called heterozygous. So an individual that is HbA HbA is homozygous normal beta chain; an individual that is HbA HbS is heterozygous; and an individual that is HbS HbS is homozygous abnormal beta chain. It is the homozygous HbS individuals that contain sickle-shaped blood cells.
Mechanism of the disease
In the capillaries (microscopic blood vessels that directly exchange oxygen with the tissues), erythrocytes can be subjected to low oxygen tension after they lose their oxygen to the surrounding tissues. In this low oxygen situation, the abnormal hemoglobin molecules of HbS HbS individuals tend to polymerize (join together), forming stiff, tubular fibers which ultimately distort the shape of the entire erythrocyte, giving it the characteristic “sickle” shape. These sickled cells have a number of effects on the body via two processes.
- Sickled cells are less able to enter and move through the capillaries: Once in the capillaries, they clog capillary flow and cause small blood clots. Reduced blood flow results in reduced oxygen availability to the tissues. Reduced oxygen supply results in tissue death and damage to vital organs (e.g., the heart, liver and spleen).
- Sickled blood cells have a shorter lifespan than normal red blood cells: Reduced lifespan of erythrocytes places a greater demand on the bone marrow to make new red blood cells and on the spleen to break down dead erythrocytes. Increased demand on the bone marrow results in severe pain in the long bones and joints. Individuals suffering from sickle cell anemia are frequently ill and generally have a considerably reduced lifespan. These individuals are said to have sickle cell disease.
Heterozygous individuals (HbA HbS) are said to be carriers for sickle cell anemia. Note that this is a specific term and is not the same thing as sickle cell anemia—heterozygotes do not have the disease themselves but their children may inherit the condition. Carriers have no anemia, have good health (as do HbA HbA individuals), and their erythrocytes maintain normal shape in the blood. In other words, they are phenotypically normal under most conditions, and probably do not know that they “carry” the HbS allele. However, if heterozygotes are exposed to conditions of low oxygen levels (such as strenuous activity at high altitudes), some of their erythrocytes do sickle. Red blood cells in blood samples of heterozygotes subjected to greatly reduced oxygen tension in the laboratory also sickle.
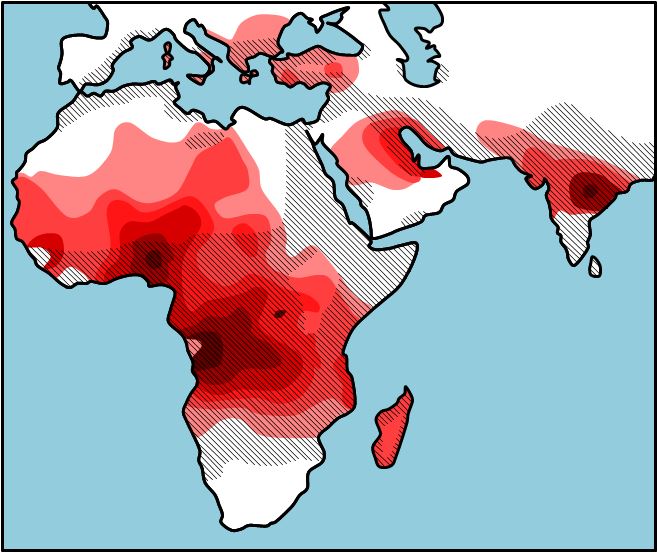
Why is sickle cell anemia most prevalent in people with origins in Central Africa and the Mediterranean? If you look at Figure 2, you will see the occurrence of sickle cell anemia overlaps with the pervasiveness of malaria. This seems odd, but those individuals who are heterozygous (HbA HbS) for the sickle cell allele are less likely to contract and die from malaria then those who are homozygous (HbA HbA). The HbS polypeptide that is produced by the heterozygous individual stops the organism (Plasmodium) that causes malaria from invading the red blood cells. So, in areas where malaria is common there is selection pressure for the HbS allele, and the HbS allele occurs in a higher frequency because the those who have one copy of the HbS allele will live longer and have more children. In areas where malaria is not common, there is selection pressure against the HbS allele, and the HbS allele occurs in a lower frequency. As you will learn in a later chapter, there is an 25% chance that two carriers will have a child who is homozygous HbS HbS), and this child will pay the evolutionary price for the protection from malaria that the parents were afforded. It seems to be in this way that evolution by natural selection retains such a potentially detrimental allele in a population. The sickle cell example is only one of what is called heterozygous advantage; we have provided a number of other examples in Table 1.
|
Distribution of malaria and the frequency of sickle cell allele
|
|---|
| Recessive illness | Heterozygote advantage | Possible explanation |
|---|---|---|
| Cystic fibrosis | Protection against diarrheal diseases such as cholera | Carriers have too few functional chloride channels in intestinal cells, blocking toxin |
| G6PD Deficiency | Protection against malaria | Red blood cells inhospitable to malaria |
| Phenylketonuria (PKU) | Protection against miscarriage induced by a fungal toxin | Excess amino acid (phenylalanine) in carriers inactivates toxin |
| Tay-Sachs disease | Protection against tuberculosis | Unknown |
| Noninsulin-dependent diabetes mellitus | Protection against starvation | Tendency to gain weight protects against starvation during famine |
GLOSSARY
- induced mutation:
-
mutation that results from exposure to chemicals or environmental agents
- mutation:
-
variation in the nucleotide sequence of a genome
- mismatch repair:
-
type of repair mechanism in which mismatched bases are removed after replication
- nucleotide excision repair:
-
type of DNA repair mechanism in which the wrong base, along with a few nucleotides upstream or downstream, are removed
- proofreading:
-
function of DNA pol in which it reads the newly added base before adding the next one
- point mutation:
-
mutation that affects a single base
- silent mutation:
-
mutation that is not expressed
- spontaneous mutation:
-
mutation that takes place in the cells as a result of chemical reactions taking place naturally without exposure to any external agent
- transition substitution:
-
when a purine is replaced with a purine or a pyrimidine is replaced with another pyrimidine
- transversion substitution:
-
when a purine is replaced by a pyrimidine or a pyrimidine is replaced by a purine