
Test Page
- Page ID
- 14479
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Lecture 14: Bacteria and Archaea and origins of Eukaryotes
Cellular structure of bacteria and archaea
In this section, we will discuss the basic structural features of both bacteria and archaea. There are many structural, morphological, and physiological similarities between bacteria and archaea. As discussed in the previous section, these microbes inhabit many ecological niches and carry out a great diversity of biochemical and metabolic processes. Both bacteria and archaea lack a membrane-bound nucleus and membrane-bound organelles, which are hallmarks of eukaryotes.
While Bacteria and Archaea are separate domains, morphologically they share a number of structural features. As a result, they face similar problems, such as the transport of nutrients into the cell, the removal of waste material from the cell, and the need to respond to rapid local environmental changes. In this section, we will focus on how their common cell structure allows them to thrive in various environments and simultaneously puts constraints on them. One of the biggest constraints is related to cell size.
Although bacteria and archaea come in a variety of shapes, the most common three shapes are as follows: cocci (spherical), bacilli (rod-shaped), and spirilli (spiral-shaped) (figure below). Both bacteria and archaea are generally small compared to typical eukaryotes. For example, most bacteria tend to be on the order of 0.2 to 1.0 µm (micrometers) in diameter and 1-10 µm in length. However, there are exceptions. Epulopiscium fishelsoni is a bacillus-shaped bacterium that is typically 80 µm in diameter and 200-600 µm long. Thiomargarita namibiensis is a spherical bacterium between 100 and 750 µm in diameter and is visible to the naked eye. For comparison, a typical human neutrophil is approximately 50 µm in diameter.

Figure 1. This figure shows the three most common shapes of bacteria and archaea: (a) cocci (spherical), (b) bacilli (rod-shaped), and (c) spirilli (spiral-shaped).
A thought question:
One question that comes to mind is why are bacteria and archaea typically so small? What are the constraints that keep them microscopic? How could bacteria such as Epulopiscium fishelsoni and Thiomargarita namibiensis overcome these constraints? Think of possible explanations or hypotheses that might answer these questions. We'll explore and develop an understanding of these questions in more detail below and in class.
The bacterial and archaeal cell: common structures
Introduction to the basic cell structure
Bacteria and archaea are unicellular organisms, which lack internal membrane-bound structures that are disconnected from the plasma membrane, a phospholipid membrane that defines the boundary between the inside and outside of the cell. In bacteria and archaea, the cytoplasmic membrane also contains all membrane-bound reactions, including those related to the electron transport chain, ATP synthase, and photosynthesis. By definition, these cells lack a nucleus. Instead, their genetic material is located in a self-defined area of the cell called the nucleoid. The bacterial and archaeal chromosome is often a single covalently closed circular double-stranded DNA molecule. However, some bacteria have linear chromosomes, and some bacteria and archaea have more than one chromosome or small non-essential circular replicating elements of DNA called plasmids. Besides the nucleoid, the next common feature is the cytoplasm (or cytosol), the "aqueous," jelly-like region encompassing the internal portion of the cell. The cytoplasm is where the soluble (non-membrane-associated) reactions occur and contains the ribosomes, the protein-RNA complex where proteins are synthesized. Finally, many bacteria and archaea also have cell walls, the rigid structural feature surrounding the plasma membrane that helps provide protection and constrain the cell shape. You should learn to create a simple sketch of a general bacterial or archaeal cell from memory.

Figure 2. The features of a typical prokaryotic cell are shown.
Constraints on the bacterial and archaeal cell
One common, almost universal, feature of bacteria and archaea is that they are small, microscopic to be exact. Even the two examples given as exceptions, Epulopiscium fishelsoni and Thiomargarita namibiensis, still face the basic constraints all bacteria and archaea face; they simply found unique strategies around the problem. So what is the largest constraint when it comes to dealing with the size of bacteria and archaea? Think about what the cell must do to survive.
Some basic requirements
So what do cells have to do to survive? They need to transform energy into a usable form. This involves making ATP, maintaining an energized membrane, and maintaining productive NAD+/NADH2 ratios. Cells also need to be able to synthesize the appropriate macromolecules (proteins, lipids, polysaccharides, etc.) and other cellular structural components. To do this, they need to be able to either make the core, key precursors for more complex molecules or get them from the environment.
Diffusion and its importance to bacteria and archaea
Movement by diffusion is passive and proceeds down the concentration gradient. For compounds to move from the outside to the inside of the cell, the compound must be able to cross the phospholipid bilayer. If the concentration of a substance is lower inside the cell than outside and it has chemical properties that allow it to move across the cell membrane, that compound will energetically tend to move into the cell. While the "real" story is a bit more complex and will be discussed in more detail later, diffusion is one of the mechanisms bacteria and archaea use to aid in the transport of metabolites.
Diffusion can also be used to get rid of some waste materials. As waste products accumulate inside the cell, their concentration rises compared to that of the outside environment, and the waste product can leave the cell. Movement within the cell works the same way: compounds will move down their concentration gradient, away from where they are synthesized to places where their concentration is low and therefore may be needed. Diffusion is a random process—the ability of two different compounds or reactants for chemical reactions to interact becomes a meeting of chance. Therefore, in small, confined spaces, random interactions or collisions can occur more frequently than they can in large spaces.
The ability of a compound to diffuse depends on the viscosity of the solvent. For example, it is a lot easier for you to move around in air than in water (think about moving around underwater in a pool). Likewise, it is easier for you to swim in a pool of water than in a pool filled with peanut butter. If you put a drop of food coloring into a glass of water, it quickly diffuses until the entire glass has changed color. Now what do you think would happen if you put that same drop of food coloring into a glass of corn syrup (very viscous and sticky)? It will take a lot longer for the glass of corn syrup to change color.
The relevance of these examples is to note that the cytoplasm tends to be very viscous. It contains many proteins, metabolites, small molecules, etc. and has a viscosity more like corn syrup than water. So, diffusion in cells is slower and more limited than you might have originally expected. Therefore, if cells rely solely on diffusion to move compounds around, what do you think happens to the efficiency of these processes as cells increase in size and their internal volumes get bigger? Is there a potential problem to getting big that is related to the process of diffusion?
So how do cells get bigger?
As you've likely concluded from the discussion above, with cells that rely on diffusion to move things around the cell—like bacteria and archaea—size does matter. So how do you suppose Epulopiscium fishelsoni and Thiomargarita namibiensis got so big? Take a look at these links, and see what these bacteria look like morphologically and structurally: Epulopiscium fishelsoni and Thiomargarita namibiensis.
Based on what we have just discussed, in order for cells to get bigger, that is, for their volume to increase, intracellular transport must somehow become independent of diffusion. One of the great evolutionary leaps was the ability of cells (eukaryotic cells) to transport compounds and materials intracellularly, independent of diffusion. Compartmentalization also provided a way to localize processes to smaller organelles, which overcame another problem caused by the large size. Compartmentalization and the complex intracellular transport systems have allowed eukaryotic cells to become very large in comparison to the diffusion-limited bacterial and archaeal cells. We'll discuss specific solutions to these challenges in the following sections.
Introduction to bacterial and archaeal diversity
Perhaps bacteria may tentatively be regarded as biochemical experiments; owing to their relatively small size and rapid growth, variations must arise much more frequently than in more differentiated forms of life, and they can in addition afford to occupy more precarious positions in natural economy than larger organisms with more exacting requirements.
Marjory Stephenson, in Bacterial Metabolism, (1930)
Prokaryotes are single-celled organisms with neither a distinct nucleus with a membrane nor other organelles. They are composed of two distinct groups of organisms: Bacteria and Archaea. In recent years, the term prokaryote has fallen out of favor for many microbiologists. The reason is that while bacteria and archaea share many morphological characteristics, they represent evolutionarily distinct domains of life. The figure below shows a simple evolutionary tree with the three main domains of life: Bacteria, Archaea, and Eukarya. Some instructors in BIS2A use the term "prokaryote" when describing the morphological characteristics of the organism but use the terms "bacteria" and "archaea" when discussing the unique characteristics of these two domains of life.
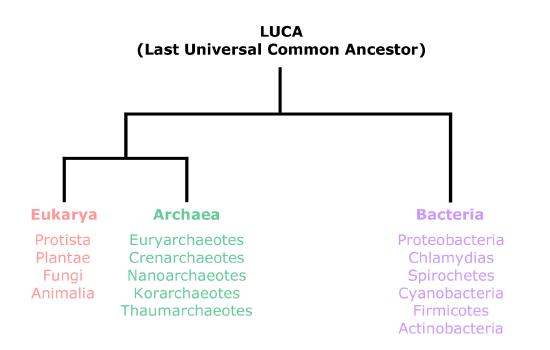
Figure 1. Although bacteria and archaea are both described as prokaryotes, they have been placed in separate domains of life. An ancestor of modern archaea is believed to have given rise to Eukarya, the third domain of life. Archaeal and bacterial phyla are shown; the evolutionary relationship between these phyla is still open to debate.
Although bacteria and archaea share many morphological, structural, and metabolic attributes, there are numerous differences between the organisms in these two clades. The most notable differences are in the chemical structure and compositions of membrane lipids (see Module 10.1), the chemical composition of the cell wall, and the makeup of the information processing machinery (e.g., replication, DNA repair, and transcription).
Bacterial and archaeal diversity
Bacteria and archaea were on Earth long before multicellular life appeared. They are ubiquitous and have highly diverse metabolic activities. This diversity allows different species within clades to inhabit every imaginable surface where there is sufficient moisture. For example, in the typical human body, bacterial cells outnumber human body cells by about ten to one. Indeed, bacteria and archaea comprise the majority of living things in all ecosystems. Certain bacterial and archaeal species can thrive in environments that are inhospitable for most other life. Bacteria and archaea, along with microbial eukaryotes, are also critical for recycling the nutrients essential for creating new biomolecules. They also drive the evolution of new ecosystems (natural or man-made).
The first inhabitants of Earth
When and where did life begin? What were the conditions on Earth when life began? Based on the fossil record, LUCA, Last Universal Common Ancestor, was the predecessor to bacteria and archaea. While we don't know what these organsims were like genetically, we do know that they had no true nucleus and were morphologically similar to bacteria and archaea. They were the first forms of life on Earth, and they existed for billions of years before plants and animals appeared. The Earth and its moon are thought to be about 4.54 billion years old. This estimate is based on evidence from radiometric dating of meteorite material, together with other substrate material from Earth and the moon. Early Earth had a very different atmosphere (contained less molecular oxygen) than it does today and was subjected to strong radiation; thus, the first organisms would have flourished in areas where they were more protected, such as in ocean depths or beneath the Earth's surface. During this time period, strong volcanic activity was common on Earth, so it is likely that these first organisms were adapted to very high temperatures. Early Earth was also bombarded with mutagenic radiation from the sun. The first organisms needed to be able to withstand all these harsh conditions.
Note:
The evolution of bacteria and archaea:
How do scientists answer questions about the evolution of bacteria and archaea? Unlike with animals, artifacts in the fossil record of bacteria and archaea offer very little information. Fossils of ancient bacteria and archaea look like tiny bubbles in rock. Some scientists turn to genetics and to the principle of the molecular clock, which holds that the more recently two species have diverged, the more similar their genes (and thus proteins) will be. Conversely, species that diverged long ago will have more genes that are dissimilar.
Scientists at the NASA Astrobiology Institute and at the European Molecular Biology Laboratory collaborated to analyze the molecular evolution of 32 specific proteins common to 72 species of bacteria. The model they derived from their data indicates that three important groups of bacteria—Actinobacteria, Deinococcus, and Cyanobacteria (which the authors call Terrabacteria)—were the first to colonize land. Deinococcus is a bacterium that is highly resistant to ionizing radiation. Cyanobacteria are photosynthesizers, while Actinobacteria are a group of very common bacteria that include species important in decomposition of organic wastes.
The timelines of divergence suggest that bacteria (members of the domain Bacteria) diverged from common ancestral species between 2.5 and 3.2 billion years ago, whereas archaea diverged earlier: between 3.1 and 4.1 billion years ago. Eukarya diverged off the Archaean line later. Furthermore, there were bacteria able to grow in the anoxic environment that existed prior to the advent of cyanobacteria (about 2.6 billion years ago). These bacteria needed to be resistance to drying and to possess compounds that protect the organism from radiation. It has been proposed that the emergence of cyanobacteria with its ability to conduct photosynthesis and produce oxygen was a key event in the evolution of life on Earth.
Microbial mats
Microbial mats (large biofilms) may represent the earliest forms of life on Earth; there is fossil evidence of their presence starting about 3.5 billion years ago. A microbial mat is a multi-layered sheet of microbes composed mostly of bacteria but that may also include archaea. Microbial mats are a few centimeters thick, and they typically grow at the interface between two materials, mostly on moist surfaces. Organisms in a microbial mat are held together by a glue-like, sticky substance that they secrete, forming an extracellular matrix. The species within the mat carry out different metabolic activities depending on their environment. As a result, microbial mats have been identified that have different textures and colors reflecting the mat composition and the metabolic activities conducted by the microorganisms that make up the mat.
The first microbial mats likely obtained their energy from chemicals found near hydrothermal vents. A hydrothermal vent is a breakage or fissure in the Earth’s surface that releases geothermally heated water. With the evolution of photosynthesis about 3 billion years ago, some organisms in microbial mats came to use a more widely available energy source—sunlight—whereas others depended on chemicals from hydrothermal vents for energy and food.

Figure 2. (a) This microbial mat, about one meter in diameter, grows over a hydrothermal vent in the Pacific Ocean in a region known as the “Pacific Ring of Fire.” The mat helps retain microbial nutrients. Chimneys, such as the one indicated by the arrow, allow gases to escape. (b) In this micrograph, bacteria within a mat are visualized using fluorescence microscopy. (credit a: modification of work by Dr. Bob Embley, NOAA PMEL, Chief Scientist; credit b: modification of work by Ricardo Murga, Rodney Donlan, CDC; scale-bar data from Matt Russell)
Stromatolites
Fossilized microbial mats represent the earliest record of life on Earth. A stromatolite is a sedimentary structure formed when minerals precipitate out of water by organisms in a microbial mat. Stromatolites form layered rocks made of carbonate or silicate. Although most stromatolites are artifacts from the past, there are places on Earth where stromatolites are still forming. For example, growing stromatolites have been found in the Anza-Borrego Desert State Park in San Diego County, California.

Figure 3. (a) These living stromatolites are located in Shark Bay, Australia. (b) These fossilized stromatolites, found in Glacier National Park, Montana, are nearly 1.5 billion years old. (credit a: Robert Young; credit b: P. Carrara, NPS).
The ancient atmosphere
Evidence indicates that during the first two billion years of Earth’s existence, the atmosphere was anoxic, meaning that there was no molecular oxygen. Therefore, only those organisms that can grow without oxygen—anaerobic organisms—were able to live. Autotrophic organisms that convert solar energy into chemical energy are called phototrophs, and they appeared within one billion years of the Earth's formation. Then, cyanobacteria, also known as blue-green algae, evolved from these simple phototrophs one billion years later. Cyanobacteria began oxygenating the atmosphere. Increased atmospheric oxygen allowed the development of more efficient O2-utilizing catabolic pathways. It also opened up the land to increased colonization, because some O2 is converted into O3 (ozone), and ozone effectively absorbs the ultraviolet light that would otherwise cause lethal mutations in DNA. Ultimately, the increase in O2 concentrations allowed the evolution of other life forms.
Bacteria and archaea are adaptable: life in moderate and extreme environments
Some organisms have developed strategies that allow them to survive harsh conditions. Bacteria and archaea thrive in a vast array of environments: some grow in conditions that would seem very normal to us, whereas others are able to thrive and grow under conditions that would kill a plant or an animal. Almost all bacteria and archaea have some form of a cell wall, a protective structure that allows them to survive in both hyper- and hypo-osmotic conditions. Some soil bacteria are able to form endospores that resist heat and drought, thereby allowing the organism to survive until more favorable conditions recur. These adaptations, along with others, allow bacteria to be the most abundant life forms in all terrestrial and aquatic ecosystems.
Some bacteria and archaea are adapted to grow under extreme conditions and are called extremophiles, meaning “lovers of extremes.” Extremophiles have been found in all kinds of environments, such as in the depths of the oceans and the earth; in hot springs, the Artic, and the Antarctic; in very dry places; in harsh chemical environments; and in high-radiation environments, just to mention a few. These organisms give us a better understanding of prokaryotic diversity and open up the possibility of finding new prokaryotic species that may lead to the discovery of new therapeutic drugs or have industrial applications. Because they have specialized adaptations that allow them to live in extreme conditions, many extremophiles cannot survive in moderate environments. There are many different groups of extremophiles. They are categorized based on the conditions in which they grow best, and several habitats are extreme in multiple ways. For example, a soda lake is both salty and alkaline, so organisms that live in a soda lake must be both alkaliphiles and halophiles. Other extremophiles, like radioresistant organisms, do not prefer an extreme environment (in this case, one with high levels of radiation) but have adapted to survive in it.
Table 1. This table lists some extremophiles and their preferred conditions.
| Extremophile Type | Conditions for Optimal Growth |
| Acidophiles | pH 3 or below |
| Alkaliphiles | pH 9 or above |
| Thermophiles | Temperature of 60–80 °C (140–176 °F) |
| Hyperthermophiles | Temperature of 80–122 °C (176–250 °F) |
| Psychrophiles | Temperature of -15 °C (5 °F) or lower |
| Halophiles | Salt concentration of at least 0.2 M |
| Osmophiles | High sugar concentration |
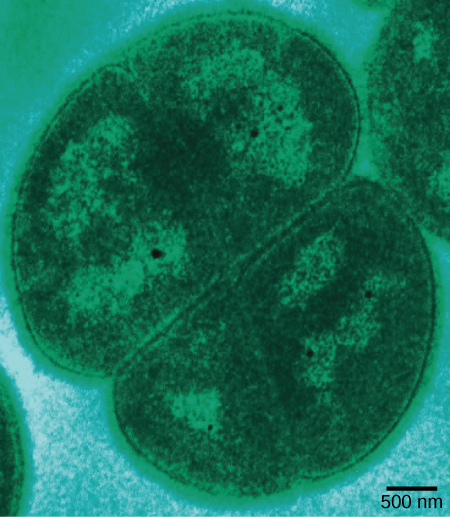
Figure 4. Deinococcus radiodurans, visualized in this false-color transmission electron micrograph, is a bacterium that can tolerate very high doses of ionizing radiation. It has developed DNA repair mechanisms that allow it to reconstruct its chromosome even if it has been broken into hundreds of pieces by radiation or heat. (credit: modification of work by Michael Daly; scale-bar data from Matt Russell)
Footnotes
1. Battistuzzi, FU, Feijao, A, and Hedges, SB. A genomic timescale of prokaryote evolution: Insights into the origin of methanogenesis, phototrophy, and the colonization of land. BioMed Central: Evolutionary Biology 4 (2004): 44, doi:10.1186/1471-2148-4-44.
Eukaryotic Cell: Structure and Function
Introduction to eukaryotic cells
By definition, eukaryotic cells are cells that contain a membrane-bound nucleus, a structural feature that is not present in bacterial or archaeal cells. In addition to the nucleus, eukaryotic cells are characterized by numerous membrane-bound organelles such as the endoplasmic reticulum, Golgi apparatus, chloroplasts, mitochondria, and others.
In previous sections, we began to consider the Design Challenge of making cells larger than a small bacterium—more precisely, growing cells to sizes at which, in the eyes of natural selection, relying on diffusion of substances for transport through a highly viscous cytosol comes with inherent functional trade-offs that offset most selective benefits of getting larger. In the lectures and readings on bacterial cell structure, we discovered some morphological features of large bacteria that allow them to effectively overcome diffusion-limited size barriers (e.g., filling the cytoplasm with a large storage vacuole maintains a small volume for metabolic activity that remains compatible with diffusion-driven transport).
As we transition our focus to eukaryotic cells, we want you to approach the study by constantly returning to the Design Challenge. We will cover a large number of subcellular structures that are unique to eukaryotes, and you will certainly be expected to know the names of these structures or organelles, to associate them with one or more "functions", and to identify them on a canonical cartoon representation of a eukaryotic cell. This memorization exercise is necessary but not sufficient. We will also ask you to start thinking a bit deeper about some of the functional and evolutionary costs and benefits (trade-offs) of both evolving eukaryotic cells and various eukaryotic organelles, as well as how a eukaryotic cell might coordinate the functions of different organelles.
Your instructors will, of course, propose some functional hypotheses for you to consider that address these broader points. Our hypotheses may sometimes come in the form of statements like, "Thing A exists because of rationale B." To be completely honest, however, in many cases, we don't actually know all of the selective pressures that led to the creation or maintenance of certain cellular structures, and the likelihood that one explanation will fit all cases is slim in biology. The causal linkage/relationship implied by the use of terms like "because" should be treated as good hypotheses rather than objective, concrete, undisputed, factual knowledge. We want you to understand these hypotheses and to be able to discuss the ideas presented in class, but we also want you to indulge your own curiosity and to begin thinking critically about these ideas yourself. Try using the Design Challenge rubric to explore some of your ideas. In the following, we will try to seed questions to encourage this activity.

Figure 1. These figures show the major organelles and other cell components of (a) a typical animal cell and (b) a typical eukaryotic plant cell. The plant cell has a cell wall, chloroplasts, plastids, and a central vacuole—structures not found in animal cells. Plant cells do not have lysosomes or centrosomes.
The plasma membrane
Like bacteria and archaea, eukaryotic cells have a plasma membrane, a phospholipid bilayer with embedded proteins that separates the internal contents of the cell from its surrounding environment. The plasma membrane controls the passage of organic molecules, ions, water, and oxygen into and out of the cell. Wastes (such as carbon dioxide and ammonia) also leave the cell by passing through the plasma membrane, usually with some help of protein transporters.

Figure 2. The eukaryotic plasma membrane is a phospholipid bilayer with proteins and cholesterol embedded in it.
As discussed in the context of bacterial cell membranes, the plasma membranes of eukaryotic cells may also adopt unique conformations. For instance, the plasma membrane of cells that, in multicellular organisms, specialize in absorption are often folded into fingerlike projections called microvilli (singular = microvillus); (see figure below). The "folding" of the membrane into microvilli effectively increases the surface area for absorption while minimally impacting the cytosolic volume. Such cells can be found lining the small intestine, the organ that absorbs nutrients from digested food.
An aside: People with celiac disease have an immune response to gluten, a protein found in wheat, barley, and rye. The immune response damages microvilli. As a consequence, afflicted individuals have an impaired ability to absorb nutrients. This can lead to malnutrition, cramping, and diarrhea.

Figure 3. Microvilli, shown here as they appear on cells lining the small intestine, increase the surface area available for absorption. These microvilli are only found on the area of the plasma membrane that faces the cavity from which substances will be absorbed. Credit: "micrograph", modification of work by Louisa Howard
The cytoplasm
The cytoplasm refers to the entire region of a cell between the plasma membrane and the nuclear envelope. It is composed of organelles suspended in the gel-like cytosol, the cytoskeleton, and various chemicals (see figure below). Even though the cytoplasm consists of 70 to 80 percent water, it nevertheless has a semisolid consistency. It is crowded in there. Proteins, simple sugars, polysaccharides, amino acids, nucleic acids, fatty acids, ions and many other water-soluble molecules are all competing for space and water.
The nucleus
Typically, the nucleus is the most prominent organelle in a cell (see figure below) when viewed through a microscope. The nucleus (plural = nuclei) houses the cell’s DNA. Let’s look at it in more detail.

Figure 4. The nucleus stores chromatin (DNA plus proteins) in a gel-like substance called the nucleoplasm. The nucleolus is a condensed region of chromatin where ribosome synthesis occurs. The boundary of the nucleus is called the nuclear envelope. It consists of two phospholipid bilayers: an outer membrane and an inner membrane. The nuclear membrane is continuous with the endoplasmic reticulum. Nuclear pores allow substances to enter and exit the nucleus.
The nuclear envelope
The nuclear envelope, a structure that constitutes the outermost boundary of the nucleus, is a double-membrane—both the inner and outer membranes of the nuclear envelope are phospholipid bilayers. The nuclear envelope is also punctuated with protein-based pores that control the passage of ions, molecules, and RNA between the nucleoplasm and cytoplasm. The nucleoplasm is the semisolid fluid inside the nucleus where we find the chromatin and the nucleolus, a condensed region of chromatin where ribosome synthesis occurs.
Chromatin and chromosomes
To understand chromatin, it is helpful to first consider chromosomes. Chromosomes are structures within the nucleus that are made up of DNA, the hereditary material. You may remember that in bacteria and archaea, DNA is typically organized into one or more circular chromosome(s). In eukaryotes, chromosomes are linear structures. Every eukaryotic species has a specific number of chromosomes in the nuclei of its cells. In humans, for example, the chromosome number is 23, while in fruit flies, it is 4.
Chromosomes are only clearly visible and distinguishable from one another by visible optical microscopy when the cell is preparing to divide and the DNA is tightly packed by proteins into easily distinguishable shapes. When the cell is in the growth and maintenance phases of its life cycle, numerous proteins are still associated with the nucleic acids, but the DNA strands more closely resemble an unwound, jumbled bunch of threads. The term chromatin is used to describe chromosomes (the protein-DNA complexes) when they are both condensed and decondensed.

Figure 5. (a) This image shows various levels of the organization of chromatin (DNA and protein). (b) This image shows paired chromosomes. Credit (b): modification of work by NIH; scale-bar data from Matt Russell
The nucleolus
Some chromosomes have sections of DNA that encode ribosomal RNA. A darkly staining area within the nucleus called the nucleolus (plural = nucleoli) aggregates the ribosomal RNA with associated proteins to assemble the ribosomal subunits that are then transported out to the cytoplasm through the pores in the nuclear envelope.
Note: possible discussion
Discuss amongst yourselves. Use the Design Challenge rubric to consider the nucleus in more detail. What "problems" does an organelle like the nucleus solve? What are some of the qualities of a nucleus that may be responsible for ensuring its evolutionary success? What are some of the trade-offs of evolving and maintaining a nucleus? (Every benefit has some cost; can you list both?) Remember, there may be some well-established hypotheses (and it is good to mention these), but the point of the exercise here is for you to think critically and to critically discuss these ideas using your collective "smarts".
Ribosomes
Ribosomes are the cellular structures responsible for protein synthesis. When viewed through an electron microscope, ribosomes appear either as clusters (polyribosomes) or single, tiny dots that float freely in the cytoplasm. They may be attached to the cytoplasmic side of the plasma membrane or the cytoplasmic side of the endoplasmic reticulum and the outer membrane of the nuclear envelope (cartoon of cell above).
Electron microscopy has shown us that ribosomes, which are large complexes of protein and RNA, consist of two subunits, aptly called large and small (figure below). Ribosomes receive their "instructions" for protein synthesis from the nucleus, where the DNA is transcribed into messenger RNA (mRNA). The mRNA travels to the ribosomes, which translate the code provided by the sequence of the nitrogenous bases in the mRNA into a specific order of amino acids in a protein. This is covered in greater detail in the section covering the process of translation.

Figure 6. Ribosomes are made up of a large subunit (top) and a small subunit (bottom). During protein synthesis, ribosomes assemble amino acids into proteins.
Mitochondria
Mitochondria (singular = mitochondrion) are often called the “powerhouses” or “energy factories” of a cell because they are the primary site of metabolic respiration in eukaryotes. Depending on the species and the type of mitochondria found in those cells, the respiratory pathways may be anaerobic or aerobic. By definition, when respiration is aerobic, the terminal electron is oxygen; when respiration is anaerobic, a compound other than oxygen functions as the terminal electron acceptor. In either case, the result of these respiratory processes is the production of ATP via oxidative phosphorylation, hence the use of terms "powerhouse" and/or "energy factory" to describe this organelle. Nearly all mitochondria also possess a small genome that encodes genes whose functions are typically restricted to the mitochondrion.
In some cases, the number of mitochondria per cell is tunable, depending, typically, on energy demand. It is for instance possible muscle cells that are used—that by extension have a higher demand for ATP—may often be found to have a significantly higher number of mitochondria than cells that do not have a high energy load.
The structure of the mitochondria can vary significantly depending on the organism and the state of the cell cycle which one is observing. The typical textbook image, however, depicts mitochondria as oval-shaped organelles with a double inner and outer membrane (see figure below); learn to recognize this generic representation. Both the inner and outer membranes are phospholipid bilayers embedded with proteins that mediate transport across them and catalyze various other biochemical reactions. The inner membrane layer has folds called cristae that increase the surface area into which respiratory chain proteins can be embedded. The region within the cristae is called the mitochondrial matrix and contains—among other things—enzymes of the TCA cycle. During respiration, protons are pumped by respiratory chain complexes from the matrix into a region known as the intermembrane space (between the inner and outer membranes).

Figure 7. This electron micrograph shows a mitochondrion as viewed with a transmission electron microscope. This organelle has an outer membrane and an inner membrane. The inner membrane contains folds, called cristae, which increase its surface area. The space between the two membranes is called the intermembrane space, and the space inside the inner membrane is called the mitochondrial matrix. ATP synthesis takes place on the inner membrane. Credit: modification of work by Matthew Britton; scale-bar data from Matt Russell
Note: possible discussion
Discuss: Processes like glycolysis, lipid biosynthesis, and nucleotide biosynthesis all have compounds that feed into the TCA cycle—some of which occurs in the mitochondria. What are some of the functional challenges associated with coordinating processes that have a common set of molecules if the enzymes are sequestered into different cellular compartments?
Peroxisomes
Peroxisomes are small, round organelles enclosed by single membranes. These organelles carry out redox reactions that oxidize and break down fatty acids and amino acids. They also help to detoxify many toxins that may enter the body. Many of these redox reactions release hydrogen peroxide, H2O2, which would be damaging to cells; however, when these reactions are confined to peroxisomes, enzymes safely break down the H2O2 into oxygen and water. For example, alcohol is detoxified by peroxisomes in liver cells. Glyoxysomes, which are specialized peroxisomes in plants, are responsible for converting stored fats into sugars.
Vesicles and vacuoles
Vesicles and vacuoles are membrane-bound sacs that function in storage and transport. Other than the fact that vacuoles are somewhat larger than vesicles, there is a very subtle distinction between them: the membranes of vesicles can fuse with either the plasma membrane or other membrane systems within the cell. Additionally, some agents such as enzymes within plant vacuoles break down macromolecules. The membrane of a vacuole does not fuse with the membranes of other cellular components.
Animal cells versus plant cells
At this point, you know that each eukaryotic cell has a plasma membrane, cytoplasm, a nucleus, ribosomes, mitochondria, peroxisomes, and in some, vacuoles. There are some striking differences between animal and plant cells worth noting. Here is a brief list of differences that we want you to be familiar with and a slightly expanded description below:
1. While all eukaryotic cells use microtubule and motor protein the based mechanisms to segregate chromosomes during cell division, the structures used to organize these microtubules differ in plants versus animal and yeast cells. Animal and yeast cells organize and anchor their microtubules into structures called microtubule organizing centers (MTOCs). These structures are composed of structures called centrioles that are composed largely of α-tubulin, β-tubulin, and other proteins. Two centrioles organize into a structure called a centrosome. By contrast, in plants, while microtubules also organize into discrete bundles, there are no conspicuous structures similar to the MTOCs seen in animal and yeast cells. Rather, depending on the organism, it appears that there can be several places where these bundles of microtubules can nucleate from places called acentriolar (without centriole) microtubule organizing centers. A third type of tubulin, γ-tubulin, appears to be implicated, but our knowledge of the precise mechanisms used by plants to organize microtubule spindles is still spotty.
2. Animal cells typically have organelles called lysosomes responsible for degradation of biomolecules. Some plant cells contain functionally similar degradative organelles, but there is a debate as to how they should be named. Some plant biologists call these organelles lysosomes while others lump them into the general category of plastids and do not give them a specific name.
3. Plant cells have a cell wall, chloroplasts and other specialized plastids, and a large central vacuole, whereas animal cells do not.
The centrosome
The centrosome is a microtubule-organizing center found near the nuclei of animal cells. It contains a pair of centrioles, two structures that lie perpendicular to eachother (see figure below). Each centriole is a cylinder of nine triplets of microtubules.

Figure 8. The centrosome consists of two centrioles that lie at right angles to each other. Each centriole is a cylinder made up of nine triplets of microtubules. Nontubulin proteins (indicated by the green lines) hold the microtubule triplets together.
The centrosome (the organelle where all microtubules originate in animal and yeast) replicates itself before a cell divides, and the centrioles appear to have some role in pulling the duplicated chromosomes to opposite ends of the dividing cell. However, the exact function of the centrioles in cell division remains unclear, as cells that have had their centrosome removed can still divide, and plant cells, which lack centrosomes, are capable of cell division.
Lysosomes
Animal cells have another set of organelles not found in plant cells: lysosomes. Colloquially, the lysosomes are sometimes called the cell’s “garbage disposal”. Enzymes within the lysosomes aid the breakdown of proteins, polysaccharides, lipids, nucleic acids, and even "worn-out" organelles. These enzymes are active at a much lower pH than that of the cytoplasm. Therefore, the pH within lysosomes is more acidic than the pH of the cytoplasm. In plant cells, many of the same digestive processes take place in vacuoles.
The cell wall
If you examine the diagram above depicting plant and animal cells, you will see in the diagram of a plant cell a structure external to the plasma membrane called the cell wall. The cell wall is a rigid covering that protects the cell, provides structural support, and gives shape to the cell. Fungal and protistan cells also have cell walls. While the chief component of bacterial cell walls is peptidoglycan, the major organic molecule in the plant cell wall is cellulose (see structure below), a polysaccharide made up of glucose subunits.

Figure 9. Cellulose is a long chain of β-glucose molecules connected by a 1-4 linkage. The dashed lines at each end of the figure indicate a series of many more glucose units. The size of the page makes it impossible to portray an entire cellulose molecule.
Chloroplasts
Chloroplasts are plant cell organelles that carry out photosynthesis. Like the mitochondria, chloroplasts have their own DNA and ribosomes, but chloroplasts have an entirely different function.
Like mitochondria, chloroplasts have outer and inner membranes, but within the space enclosed by a chloroplast’s inner membrane is a set of interconnected and stacked fluid-filled membrane sacs called thylakoids (figure below). Each stack of thylakoids is called a granum (plural = grana). The fluid enclosed by the inner membrane that surrounds the grana is called the stroma.

Figure 10. The chloroplast has an outer membrane, an inner membrane, and membrane structures called thylakoids that are stacked into grana. The space inside the thylakoid membranes is called the thylakoid space. The light harvesting reactions take place in the thylakoid membranes, and the synthesis of sugar takes place in the fluid inside the inner membrane, which is called the stroma. Chloroplasts also have their own genome, which is contained on a single circular chromosome.
The chloroplasts contain a green pigment called chlorophyll, which captures the light energy that drives the reactions of photosynthesis. Like plant cells, photosynthetic protists also have chloroplasts. Some bacteria perform photosynthesis, but their chlorophyll is not relegated to an organelle.
Evolution connection
Endosymbiosis
We have mentioned that both mitochondria and chloroplasts contain DNA and ribosomes. Have you wondered why? Strong evidence points to endosymbiosis as the explanation.
Symbiosis is a relationship in which organisms from two separate species depend on each other for their survival. Endosymbiosis (endo- = “within”) is a mutually beneficial relationship in which one organism lives inside the other. Endosymbiotic relationships abound in nature. For instance, some microbes that live in our digestive tracks produce vitamin K. The relationship between these microbes and us (their hosts) is said to be mutually beneficial or symbiotic. The relationship is beneficial for us because we are unable to synthesize vitamin K; the microbes do it for us instead. The relationship is also beneficial for the microbes because they receive abundant food from the environment of the large intestine, and they are protected both from other organisms and from drying out.
Scientists have long noticed that bacteria, mitochondria, and chloroplasts are similar in size. We also know that bacteria have DNA and ribosomes, just as mitochondria and chloroplasts do. Scientists believe that host cells and bacteria formed an endosymbiotic relationship when the host cells ingested both aerobic and autotrophic bacteria (cyanobacteria) but did not destroy them. Through many millions of years of evolution, these ingested bacteria became more specialized in their functions, with the aerobic bacteria becoming mitochondria and the autotrophic bacteria becoming chloroplasts. There will be more on this later in the reading.
The central vacuole
Previously, we mentioned vacuoles as essential components of plant cells. If you look at the cartoon figure of the plant cell, you will see that it depicts a large central vacuole that occupies most of the area of the cell. The central vacuole plays a key role in regulating the cell’s concentration of water in changing environmental conditions.
Silly vacuole factoid: Have you ever noticed that if you forget to water a plant for a few days, it wilts? That’s because as the water concentration in the soil becomes lower than the water concentration in the plant, water moves out of the central vacuoles and cytoplasm. As the central vacuole shrinks, it leaves the cell wall unsupported. This loss of support to the cell walls of plant cells results in the wilted appearance of the plant.
The central vacuole also supports the expansion of the cell. When the central vacuole holds more water, the cell gets larger without having to invest a lot of energy in synthesizing new cytoplasm.
Eukaryotes
Living things fall into three large groups: Archaea, Bacteria, and Eukarya. The first two groups include non-nucleated cells, and the third contains all eukaryotes. A relatively sparse fossil record is available to help us discern what the first members of each of these lineages looked like, so it is possible that all the events that led up to the last common ancestor of extant eukaryotes will remain unknown. However, comparative biology of extant organisms and the limited fossil record provide some insights into the history of Eukarya.
The earliest fossils found appear to be bacteria, most likely cyanobacteria. They are about 3.5 billion years old and are recognizable because of their relatively complex structure and, for bacteria, relatively large cells. Most other bacteria and archaea have small cells, 1 or 2 µm in size, and would be difficult to pick out as fossils. Most living eukaryotes have cells measuring 10 µm or greater. Structures this size, which might be fossils, appear in the geological record about 2.1 billion years ago.
Characteristics of eukaryotes
Data from these fossils have led biologists to the conclusion that living eukaryotes are all descendants of a single common ancestor. Mapping the characteristics found in all major groups of eukaryotes reveals that the following characteristics must have been present in the last common ancestor, because these characteristics are present in at least some of the members of each major lineage.
- Cells with nuclei surrounded by a nuclear envelope with nuclear pores. This is the single characteristic that is both necessary and sufficient to define an organism as a eukaryote. All extant eukaryotes have cells with nuclei.
- Mitochondria. Some extant eukaryotes have very reduced remnants of mitochondria in their cells, whereas other members of their lineages have “typical” mitochondria.
- A cytoskeleton containing the structural and motility components called actin microfilaments and microtubules. All extant eukaryotes have these cytoskeletal elements.
- Flagella and cilia, organelles associated with cell motility. Some extant eukaryotes lack flagella and/or cilia, but they are descended from ancestors that possessed them.
- Chromosomes, each consisting of a linear DNA molecule coiled around basic (alkaline) proteins called histones. The few eukaryotes with chromosomes lacking histones clearly evolved from ancestors that had them.
- Mitosis, a process of nuclear division wherein replicated chromosomes are divided and separated using elements of the cytoskeleton. Mitosis is universally present in eukaryotes.
- Sex, a process of genetic recombination unique to eukaryotes in which diploid nuclei at one stage of the life cycle undergo meiosis to yield haploid nuclei and subsequent karyogamy, a stage where two haploid nuclei fuse together to create a diploid zygote nucleus.
- Members of all major lineages have cell walls, and it might be reasonable to conclude that the last common ancestor could make cell walls during some stage of its life cycle. However, not enough is known about eukaryotes’ cell walls and their development to know how much homology exists among them. If the last common ancestor could make cell walls, it is clear that this ability must have been lost in many groups.
Endosymbiosis and the evolution of eukaryotes
In order to understand eukaryotic organisms fully, it is necessary to understand that all extant eukaryotes are descendants of a chimeric organism that was a composite of a host cell and the cell(s) of an alpha-proteobacterium that “took up residence” inside it. This major theme in the origin of eukaryotes is known as endosymbiosis, one cell engulfing another such that the engulfed cell survives and both cells benefit. Over many generations, a symbiotic relationship can result in two organisms that depend on each other so completely that neither could survive on its own. Endosymbiotic events likely contributed to the origin of the last common ancestor of today’s eukaryotes and to later diversification in certain lineages of eukaryotes. Before explaining this further, it is necessary to consider metabolism in bacteria and archaea.
Bacterial and archaeal metabolism
Many important metabolic processes arose in bacteria and archaea, and some of these, such as nitrogen fixation, are never found in eukaryotes. The process of aerobic respiration is found in all major lineages of eukaryotes, and it is localized in the mitochondria. Aerobic respiration is also found in many lineages of bacteria and archaea, but it is not present in all of them, and many forms of evidence suggest that such anaerobic microbes never carried out aerobic respiration nor did their ancestors.
While today’s atmosphere is about one-fifth molecular oxygen (O2), geological evidence shows that it originally lacked O2. Without oxygen, aerobic respiration would not be expected, and living things would have relied on fermentation instead. At some point around 3.5 billion years ago, some bacteria and archaea began using energy from sunlight to power anabolic processes that reduce carbon dioxide to form organic compounds. That is, they evolved the ability to photosynthesize. Hydrogen, derived from various sources, was captured using light-powered reactions to reduce fixed carbon dioxide in the Calvin cycle. The group of Gram-negative bacteria that gave rise to cyanobacteria used water as the hydrogen source and released O2 as a waste product.
Eventually, the amount of photosynthetic oxygen built up in some environments to levels that posed a risk to living organisms, since it can damage many organic compounds. Various metabolic processes evolved that protected organisms from oxygen; one of which, aerobic respiration, also generated high levels of ATP. It became widely present among microbes, including in a group we now call alpha-proteobacteria. Organisms that did not acquire aerobic respiration had to remain in oxygen-free environments. Originally, oxygen-rich environments were likely localized around places where cyanobacteria were active, but by about 2 billion years ago, geological evidence shows that oxygen was building up to higher concentrations in the atmosphere. Oxygen levels similar to today’s levels only arose within the last 700 million years.
Recall that the first fossils that we believe to be eukaryotes are about 2 billion years old, so they appeared as oxygen levels were increasing. Also, recall that all extant eukaryotes descended from an ancestor with mitochondria. These organelles were first observed by light microscopists in the late 1800s, where they appeared to be somewhat worm-shaped structures that seemed to be moving around in the cell. Some early observers suggested that they might be bacteria living inside host cells, but these hypotheses remained unknown or rejected in most scientific communities.
Endosymbiotic theory
As cell biology developed in the twentieth century, it became clear that mitochondria were the organelles responsible for producing ATP using aerobic respiration. In the 1960s, American biologist Lynn Margulis developed endosymbiotic theory, which states that eukaryotes may have been a product of one cell engulfing another (one living within another) and evolving over time until the separate cells were no longer recognizable as such. In 1967, Margulis introduced new work on the theory and substantiated her findings through microbiological evidence. Although Margulis’ work initially was met with resistance, this once-revolutionary hypothesis is now widely (but not completely) accepted, with work progressing on uncovering the steps involved in this evolutionary process and the key players involved. Much still remains to be discovered about the origins of the cells that now make up the cells in all living eukaryotes.
Broadly, it has become clear that many of our nuclear genes and the molecular machinery responsible for replication and expression appear closely related to those in Archaea. On the other hand, the metabolic organelles and genes responsible for many energy-harvesting processes had their origins in bacteria. Much remains to be clarified about how this relationship occurred; this continues to be an exciting field of discovery in biology. For instance, it is not known whether the endosymbiotic event that led to mitochondria occurred before or after the host cell had a nucleus. Such organisms would be among the extinct precursors of the last common ancestor of eukaryotes.
Mitochondria
One of the major features distinguishing bacteria and archaea from eukaryotes is the presence of mitochondria. Eukaryotic cells may contain anywhere from one to several thousand mitochondria, depending on the cell’s level of energy consumption. Each mitochondrion measures 1 to 10 or greater micrometers in length and exists in the cell as an organelle that can be ovoid, worm-shaped, or intricately branched. Mitochondria arise from the division of existing mitochondria; they may fuse together; and they may be moved around inside the cell by interactions with the cytoskeleton. However, mitochondria cannot survive outside the cell. As the atmosphere was oxygenated by photosynthesis, and as successful aerobic microbes evolved, evidence suggests that an ancestral cell with some membrane compartmentalization engulfed a free-living aerobic bacterium, specifically an alpha-proteobacterium, thereby giving the host cell the ability to use oxygen to release energy stored in nutrients. Alpha-proteobacteria are a large group of bacteria that includes species symbiotic with plants, disease organisms that can infect humans via ticks, and many free-living species that use light for energy. Several lines of evidence support that mitochondria are derived from this endosymbiotic event. Most mitochondria are shaped like alpha-proteobacteria and are surrounded by two membranes, which would result when one membrane-bound organism is engulfed into a vacuole by another membrane-bound organism. The mitochondrial inner membrane is extensive and involves substantial infoldings called cristae that resemble the textured, outer surface of alpha-proteobacteria. The matrix and inner membrane are rich with the enzymes necessary for aerobic respiration.