
BIS 2A Ireland Lecture 2
- Page ID
- 11589
What is Life?
The question "What is life" has been discussed by people throughout the ages. Some people respond using metaphysical terms; others respond based on their belief system. In BIS2A, we define "life" based on our observations concerning its characteristics using scientific terms. However, it is important that you realize that not all scientific fields use the same definition. As biology becomes a more interdisciplinary science, it is important for students in the Life Sciences to appreciate this issue and understand the reason for the different definitions in different scientific fields.
possible discussion: Developing your own definition
Take the time to write down your own definition of the word "life." It can be in the form of a simple sentence, a list of bullet points, a paragraph, or a series of pictures. Then, look-up the definition of "life" on the internet. (One way would be to insert "define: Life" into the search bar.) You will notice that there are lots of definitions. How do these definitions compare with your own definition? After reading these definitions, did you find that you needed to expand your own definition?
possible discussion: Life's Working Definition: Does It Work?
NASA has been trying to develop a working definition of "life" for years. After all, how will the astrobiologists know that they have discovered a new "life" from if they don't have a definition for "life." The difficulty of coming up with a specific definition for "life" was discussed in a feature article in NASA's Astrobiology Magazine entitled Life's Working Definition: Does It Work? (https://www.nasa.gov/vision/universe...efinition.html).
The authors state that "Comparing the semantic task to the ancient Hindu story of identifying an elephant by having each of six blind men touch only the tail, the trunk, or the leg, what answer a biologist might give can differ dramatically from the answer given by a theoretical physicist." What point do you think the authors are trying to make with this statement? Do you agree that the semantic task is really this difficult?
Cellular structure of bacteria and archaea
In this section, we will discuss the basic structural features of both bacteria and archaea. There are many structural, morphological, and physiological similarities between bacteria and archaea. As discussed in the previous section, these microbes inhabit many ecological niches and carry out a great diversity of biochemical and metabolic processes. Both bacteria and archaea lack a membrane-bound nucleus and membrane-bound organelles, which are hallmarks of eukaryotes.
While Bacteria and Archaea are separate domains, morphologically they share a number of structural features. As a result, they face similar problems, such as the transport of nutrients into the cell, the removal of waste material from the cell, and the need to respond to rapid local environmental changes. In this section, we will focus on how their common cell structure allows them to thrive in various environments and simultaneously puts constraints on them. One of the biggest constraints is related to cell size.
Although bacteria and archaea come in a variety of shapes, the most common three shapes are as follows: cocci (spherical), bacilli (rod-shaped), and spirilli (spiral-shaped) (figure below). Both bacteria and archaea are generally small compared to typical eukaryotes. For example, most bacteria tend to be on the order of 0.2 to 1.0 µm (micrometers) in diameter and 1-10 µm in length. However, there are exceptions. Epulopiscium fishelsoni is a bacillus-shaped bacterium that is typically 80 µm in diameter and 200-600 µm long. Thiomargarita namibiensis is a spherical bacterium between 100 and 750 µm in diameter and is visible to the naked eye. For comparison, a typical human neutrophil is approximately 50 µm in diameter.

Figure 1. This figure shows the three most common shapes of bacteria and archaea: (a) cocci (spherical), (b) bacilli (rod-shaped), and (c) spirilli (spiral-shaped).
A thought question:
One question that comes to mind is why are bacteria and archaea typically so small? What are the constraints that keep them microscopic? How could bacteria such as Epulopiscium fishelsoni and Thiomargarita namibiensis overcome these constraints? Think of possible explanations or hypotheses that might answer these questions. We'll explore and develop an understanding of these questions in more detail below and in class.
The bacterial and archaeal cell: common structures
Introduction to the basic cell structure
Bacteria and archaea are unicellular organisms, which lack internal membrane-bound structures that are disconnected from the plasma membrane, a phospholipid membrane that defines the boundary between the inside and outside of the cell. In bacteria and archaea, the cytoplasmic membrane also contains all membrane-bound reactions, including those related to the electron transport chain, ATP synthase, and photosynthesis. By definition, these cells lack a nucleus. Instead, their genetic material is located in a self-defined area of the cell called the nucleoid. The bacterial and archaeal chromosome is often a single covalently closed circular double-stranded DNA molecule. However, some bacteria have linear chromosomes, and some bacteria and archaea have more than one chromosome or small non-essential circular replicating elements of DNA called plasmids. Besides the nucleoid, the next common feature is the cytoplasm (or cytosol), the "aqueous," jelly-like region encompassing the internal portion of the cell. The cytoplasm is where the soluble (non-membrane-associated) reactions occur and contains the ribosomes, the protein-RNA complex where proteins are synthesized. Finally, many bacteria and archaea also have cell walls, the rigid structural feature surrounding the plasma membrane that helps provide protection and constrain the cell shape. You should learn to create a simple sketch of a general bacterial or archaeal cell from memory.

Figure 2. The features of a typical prokaryotic cell are shown.
Constraints on the bacterial and archaeal cell
One common, almost universal, feature of bacteria and archaea is that they are small, microscopic to be exact. Even the two examples given as exceptions, Epulopiscium fishelsoni and Thiomargarita namibiensis, still face the basic constraints all bacteria and archaea face; they simply found unique strategies around the problem. So what is the largest constraint when it comes to dealing with the size of bacteria and archaea? Think about what the cell must do to survive.
Some basic requirements
So what do cells have to do to survive? They need to transform energy into a usable form. This involves making ATP, maintaining an energized membrane, and maintaining productive NAD+/NADH2 ratios. Cells also need to be able to synthesize the appropriate macromolecules (proteins, lipids, polysaccharides, etc.) and other cellular structural components. To do this, they need to be able to either make the core, key precursors for more complex molecules or get them from the environment.
Diffusion and its importance to bacteria and archaea
Movement by diffusion is passive and proceeds down the concentration gradient. For compounds to move from the outside to the inside of the cell, the compound must be able to cross the phospholipid bilayer. If the concentration of a substance is lower inside the cell than outside and it has chemical properties that allow it to move across the cell membrane, that compound will energetically tend to move into the cell. While the "real" story is a bit more complex and will be discussed in more detail later, diffusion is one of the mechanisms bacteria and archaea use to aid in the transport of metabolites.
Diffusion can also be used to get rid of some waste materials. As waste products accumulate inside the cell, their concentration rises compared to that of the outside environment, and the waste product can leave the cell. Movement within the cell works the same way: compounds will move down their concentration gradient, away from where they are synthesized to places where their concentration is low and therefore may be needed. Diffusion is a random process—the ability of two different compounds or reactants for chemical reactions to interact becomes a meeting of chance. Therefore, in small, confined spaces, random interactions or collisions can occur more frequently than they can in large spaces.
The ability of a compound to diffuse depends on the viscosity of the solvent. For example, it is a lot easier for you to move around in air than in water (think about moving around underwater in a pool). Likewise, it is easier for you to swim in a pool of water than in a pool filled with peanut butter. If you put a drop of food coloring into a glass of water, it quickly diffuses until the entire glass has changed color. Now what do you think would happen if you put that same drop of food coloring into a glass of corn syrup (very viscous and sticky)? It will take a lot longer for the glass of corn syrup to change color.
The relevance of these examples is to note that the cytoplasm tends to be very viscous. It contains many proteins, metabolites, small molecules, etc. and has a viscosity more like corn syrup than water. So, diffusion in cells is slower and more limited than you might have originally expected. Therefore, if cells rely solely on diffusion to move compounds around, what do you think happens to the efficiency of these processes as cells increase in size and their internal volumes get bigger? Is there a potential problem to getting big that is related to the process of diffusion?
So how do cells get bigger?
As you've likely concluded from the discussion above, with cells that rely on diffusion to move things around the cell—like bacteria and archaea—size does matter. So how do you suppose Epulopiscium fishelsoni and Thiomargarita namibiensis got so big? Take a look at these links, and see what these bacteria look like morphologically and structurally: Epulopiscium fishelsoni and Thiomargarita namibiensis.
Based on what we have just discussed, in order for cells to get bigger, that is, for their volume to increase, intracellular transport must somehow become independent of diffusion. One of the great evolutionary leaps was the ability of cells (eukaryotic cells) to transport compounds and materials intracellularly, independent of diffusion. Compartmentalization also provided a way to localize processes to smaller organelles, which overcame another problem caused by the large size. Compartmentalization and the complex intracellular transport systems have allowed eukaryotic cells to become very large in comparison to the diffusion-limited bacterial and archaeal cells. We'll discuss specific solutions to these challenges in the following sections.
Eukaryotic Cell: Structure and Function
Introduction to eukaryotic cells
By definition, eukaryotic cells are cells that contain a membrane-bound nucleus, a structural feature that is not present in bacterial or archaeal cells. In addition to the nucleus, eukaryotic cells are characterized by numerous membrane-bound organelles such as the endoplasmic reticulum, Golgi apparatus, chloroplasts, mitochondria, and others.
In previous sections, we began to consider the Design Challenge of making cells larger than a small bacterium—more precisely, growing cells to sizes at which, in the eyes of natural selection, relying on diffusion of substances for transport through a highly viscous cytosol comes with inherent functional trade-offs that offset most selective benefits of getting larger. In the lectures and readings on bacterial cell structure, we discovered some morphological features of large bacteria that allow them to effectively overcome diffusion-limited size barriers (e.g., filling the cytoplasm with a large storage vacuole maintains a small volume for metabolic activity that remains compatible with diffusion-driven transport).
As we transition our focus to eukaryotic cells, we want you to approach the study by constantly returning to the Design Challenge. We will cover a large number of subcellular structures that are unique to eukaryotes, and you will certainly be expected to know the names of these structures or organelles, to associate them with one or more "functions", and to identify them on a canonical cartoon representation of a eukaryotic cell. This memorization exercise is necessary but not sufficient. We will also ask you to start thinking a bit deeper about some of the functional and evolutionary costs and benefits (trade-offs) of both evolving eukaryotic cells and various eukaryotic organelles, as well as how a eukaryotic cell might coordinate the functions of different organelles.
Your instructors will, of course, propose some functional hypotheses for you to consider that address these broader points. Our hypotheses may sometimes come in the form of statements like, "Thing A exists because of rationale B." To be completely honest, however, in many cases, we don't actually know all of the selective pressures that led to the creation or maintenance of certain cellular structures, and the likelihood that one explanation will fit all cases is slim in biology. The causal linkage/relationship implied by the use of terms like "because" should be treated as good hypotheses rather than objective, concrete, undisputed, factual knowledge. We want you to understand these hypotheses and to be able to discuss the ideas presented in class, but we also want you to indulge your own curiosity and to begin thinking critically about these ideas yourself. Try using the Design Challenge rubric to explore some of your ideas. In the following, we will try to seed questions to encourage this activity.

Figure 1. These figures show the major organelles and other cell components of (a) a typical animal cell and (b) a typical eukaryotic plant cell. The plant cell has a cell wall, chloroplasts, plastids, and a central vacuole—structures not found in animal cells. Plant cells do not have lysosomes or centrosomes.
The plasma membrane
Like bacteria and archaea, eukaryotic cells have a plasma membrane, a phospholipid bilayer with embedded proteins that separates the internal contents of the cell from its surrounding environment. The plasma membrane controls the passage of organic molecules, ions, water, and oxygen into and out of the cell. Wastes (such as carbon dioxide and ammonia) also leave the cell by passing through the plasma membrane, usually with some help of protein transporters.

Figure 2. The eukaryotic plasma membrane is a phospholipid bilayer with proteins and cholesterol embedded in it.
As discussed in the context of bacterial cell membranes, the plasma membranes of eukaryotic cells may also adopt unique conformations. For instance, the plasma membrane of cells that, in multicellular organisms, specialize in absorption are often folded into fingerlike projections called microvilli (singular = microvillus); (see figure below). The "folding" of the membrane into microvilli effectively increases the surface area for absorption while minimally impacting the cytosolic volume. Such cells can be found lining the small intestine, the organ that absorbs nutrients from digested food.
An aside: People with celiac disease have an immune response to gluten, a protein found in wheat, barley, and rye. The immune response damages microvilli. As a consequence, afflicted individuals have an impaired ability to absorb nutrients. This can lead to malnutrition, cramping, and diarrhea.

Figure 3. Microvilli, shown here as they appear on cells lining the small intestine, increase the surface area available for absorption. These microvilli are only found on the area of the plasma membrane that faces the cavity from which substances will be absorbed. Credit: "micrograph", modification of work by Louisa Howard
The cytoplasm
The cytoplasm refers to the entire region of a cell between the plasma membrane and the nuclear envelope. It is composed of organelles suspended in the gel-like cytosol, the cytoskeleton, and various chemicals (see figure below). Even though the cytoplasm consists of 70 to 80 percent water, it nevertheless has a semisolid consistency. It is crowded in there. Proteins, simple sugars, polysaccharides, amino acids, nucleic acids, fatty acids, ions and many other water-soluble molecules are all competing for space and water.
The nucleus
Typically, the nucleus is the most prominent organelle in a cell (see figure below) when viewed through a microscope. The nucleus (plural = nuclei) houses the cell’s DNA. Let’s look at it in more detail.

Figure 4. The nucleus stores chromatin (DNA plus proteins) in a gel-like substance called the nucleoplasm. The nucleolus is a condensed region of chromatin where ribosome synthesis occurs. The boundary of the nucleus is called the nuclear envelope. It consists of two phospholipid bilayers: an outer membrane and an inner membrane. The nuclear membrane is continuous with the endoplasmic reticulum. Nuclear pores allow substances to enter and exit the nucleus.
The nuclear envelope
The nuclear envelope, a structure that constitutes the outermost boundary of the nucleus, is a double-membrane—both the inner and outer membranes of the nuclear envelope are phospholipid bilayers. The nuclear envelope is also punctuated with protein-based pores that control the passage of ions, molecules, and RNA between the nucleoplasm and cytoplasm. The nucleoplasm is the semisolid fluid inside the nucleus where we find the chromatin and the nucleolus, a condensed region of chromatin where ribosome synthesis occurs.
Chromatin and chromosomes
To understand chromatin, it is helpful to first consider chromosomes. Chromosomes are structures within the nucleus that are made up of DNA, the hereditary material. You may remember that in bacteria and archaea, DNA is typically organized into one or more circular chromosome(s). In eukaryotes, chromosomes are linear structures. Every eukaryotic species has a specific number of chromosomes in the nuclei of its cells. In humans, for example, the chromosome number is 23, while in fruit flies, it is 4.
Chromosomes are only clearly visible and distinguishable from one another by visible optical microscopy when the cell is preparing to divide and the DNA is tightly packed by proteins into easily distinguishable shapes. When the cell is in the growth and maintenance phases of its life cycle, numerous proteins are still associated with the nucleic acids, but the DNA strands more closely resemble an unwound, jumbled bunch of threads. The term chromatin is used to describe chromosomes (the protein-DNA complexes) when they are both condensed and decondensed.

Figure 5. (a) This image shows various levels of the organization of chromatin (DNA and protein). (b) This image shows paired chromosomes. Credit (b): modification of work by NIH; scale-bar data from Matt Russell
The nucleolus
Some chromosomes have sections of DNA that encode ribosomal RNA. A darkly staining area within the nucleus called the nucleolus (plural = nucleoli) aggregates the ribosomal RNA with associated proteins to assemble the ribosomal subunits that are then transported out to the cytoplasm through the pores in the nuclear envelope.
Note: possible discussion
Discuss amongst yourselves. Use the Design Challenge rubric to consider the nucleus in more detail. What "problems" does an organelle like the nucleus solve? What are some of the qualities of a nucleus that may be responsible for ensuring its evolutionary success? What are some of the trade-offs of evolving and maintaining a nucleus? (Every benefit has some cost; can you list both?) Remember, there may be some well-established hypotheses (and it is good to mention these), but the point of the exercise here is for you to think critically and to critically discuss these ideas using your collective "smarts".
Ribosomes
Ribosomes are the cellular structures responsible for protein synthesis. When viewed through an electron microscope, ribosomes appear either as clusters (polyribosomes) or single, tiny dots that float freely in the cytoplasm. They may be attached to the cytoplasmic side of the plasma membrane or the cytoplasmic side of the endoplasmic reticulum and the outer membrane of the nuclear envelope (cartoon of cell above).
Electron microscopy has shown us that ribosomes, which are large complexes of protein and RNA, consist of two subunits, aptly called large and small (figure below). Ribosomes receive their "instructions" for protein synthesis from the nucleus, where the DNA is transcribed into messenger RNA (mRNA). The mRNA travels to the ribosomes, which translate the code provided by the sequence of the nitrogenous bases in the mRNA into a specific order of amino acids in a protein. This is covered in greater detail in the section covering the process of translation.

Figure 6. Ribosomes are made up of a large subunit (top) and a small subunit (bottom). During protein synthesis, ribosomes assemble amino acids into proteins.
Mitochondria
Mitochondria (singular = mitochondrion) are often called the “powerhouses” or “energy factories” of a cell because they are the primary site of metabolic respiration in eukaryotes. Depending on the species and the type of mitochondria found in those cells, the respiratory pathways may be anaerobic or aerobic. By definition, when respiration is aerobic, the terminal electron is oxygen; when respiration is anaerobic, a compound other than oxygen functions as the terminal electron acceptor. In either case, the result of these respiratory processes is the production of ATP via oxidative phosphorylation, hence the use of terms "powerhouse" and/or "energy factory" to describe this organelle. Nearly all mitochondria also possess a small genome that encodes genes whose functions are typically restricted to the mitochondrion.
In some cases, the number of mitochondria per cell is tunable, depending, typically, on energy demand. It is for instance possible muscle cells that are used—that by extension have a higher demand for ATP—may often be found to have a significantly higher number of mitochondria than cells that do not have a high energy load.
The structure of the mitochondria can vary significantly depending on the organism and the state of the cell cycle which one is observing. The typical textbook image, however, depicts mitochondria as oval-shaped organelles with a double inner and outer membrane (see figure below); learn to recognize this generic representation. Both the inner and outer membranes are phospholipid bilayers embedded with proteins that mediate transport across them and catalyze various other biochemical reactions. The inner membrane layer has folds called cristae that increase the surface area into which respiratory chain proteins can be embedded. The region within the cristae is called the mitochondrial matrix and contains—among other things—enzymes of the TCA cycle. During respiration, protons are pumped by respiratory chain complexes from the matrix into a region known as the intermembrane space (between the inner and outer membranes).

Figure 7. This electron micrograph shows a mitochondrion as viewed with a transmission electron microscope. This organelle has an outer membrane and an inner membrane. The inner membrane contains folds, called cristae, which increase its surface area. The space between the two membranes is called the intermembrane space, and the space inside the inner membrane is called the mitochondrial matrix. ATP synthesis takes place on the inner membrane. Credit: modification of work by Matthew Britton; scale-bar data from Matt Russell
Note: possible discussion
Discuss: Processes like glycolysis, lipid biosynthesis, and nucleotide biosynthesis all have compounds that feed into the TCA cycle—some of which occurs in the mitochondria. What are some of the functional challenges associated with coordinating processes that have a common set of molecules if the enzymes are sequestered into different cellular compartments?
Peroxisomes
Peroxisomes are small, round organelles enclosed by single membranes. These organelles carry out redox reactions that oxidize and break down fatty acids and amino acids. They also help to detoxify many toxins that may enter the body. Many of these redox reactions release hydrogen peroxide, H2O2, which would be damaging to cells; however, when these reactions are confined to peroxisomes, enzymes safely break down the H2O2 into oxygen and water. For example, alcohol is detoxified by peroxisomes in liver cells. Glyoxysomes, which are specialized peroxisomes in plants, are responsible for converting stored fats into sugars.
Vesicles and vacuoles
Vesicles and vacuoles are membrane-bound sacs that function in storage and transport. Other than the fact that vacuoles are somewhat larger than vesicles, there is a very subtle distinction between them: the membranes of vesicles can fuse with either the plasma membrane or other membrane systems within the cell. Additionally, some agents such as enzymes within plant vacuoles break down macromolecules. The membrane of a vacuole does not fuse with the membranes of other cellular components.
Animal cells versus plant cells
At this point, you know that each eukaryotic cell has a plasma membrane, cytoplasm, a nucleus, ribosomes, mitochondria, peroxisomes, and in some, vacuoles. There are some striking differences between animal and plant cells worth noting. Here is a brief list of differences that we want you to be familiar with and a slightly expanded description below:
1. While all eukaryotic cells use microtubule and motor protein the based mechanisms to segregate chromosomes during cell division, the structures used to organize these microtubules differ in plants versus animal and yeast cells. Animal and yeast cells organize and anchor their microtubules into structures called microtubule organizing centers (MTOCs). These structures are composed of structures called centrioles that are composed largely of α-tubulin, β-tubulin, and other proteins. Two centrioles organize into a structure called a centrosome. By contrast, in plants, while microtubules also organize into discrete bundles, there are no conspicuous structures similar to the MTOCs seen in animal and yeast cells. Rather, depending on the organism, it appears that there can be several places where these bundles of microtubules can nucleate from places called acentriolar (without centriole) microtubule organizing centers. A third type of tubulin, γ-tubulin, appears to be implicated, but our knowledge of the precise mechanisms used by plants to organize microtubule spindles is still spotty.
2. Animal cells typically have organelles called lysosomes responsible for degradation of biomolecules. Some plant cells contain functionally similar degradative organelles, but there is a debate as to how they should be named. Some plant biologists call these organelles lysosomes while others lump them into the general category of plastids and do not give them a specific name.
3. Plant cells have a cell wall, chloroplasts and other specialized plastids, and a large central vacuole, whereas animal cells do not.
The centrosome
The centrosome is a microtubule-organizing center found near the nuclei of animal cells. It contains a pair of centrioles, two structures that lie perpendicular to eachother (see figure below). Each centriole is a cylinder of nine triplets of microtubules.

Figure 8. The centrosome consists of two centrioles that lie at right angles to each other. Each centriole is a cylinder made up of nine triplets of microtubules. Nontubulin proteins (indicated by the green lines) hold the microtubule triplets together.
The centrosome (the organelle where all microtubules originate in animal and yeast) replicates itself before a cell divides, and the centrioles appear to have some role in pulling the duplicated chromosomes to opposite ends of the dividing cell. However, the exact function of the centrioles in cell division remains unclear, as cells that have had their centrosome removed can still divide, and plant cells, which lack centrosomes, are capable of cell division.
Lysosomes
Animal cells have another set of organelles not found in plant cells: lysosomes. Colloquially, the lysosomes are sometimes called the cell’s “garbage disposal”. Enzymes within the lysosomes aid the breakdown of proteins, polysaccharides, lipids, nucleic acids, and even "worn-out" organelles. These enzymes are active at a much lower pH than that of the cytoplasm. Therefore, the pH within lysosomes is more acidic than the pH of the cytoplasm. In plant cells, many of the same digestive processes take place in vacuoles.
The cell wall
If you examine the diagram above depicting plant and animal cells, you will see in the diagram of a plant cell a structure external to the plasma membrane called the cell wall. The cell wall is a rigid covering that protects the cell, provides structural support, and gives shape to the cell. Fungal and protistan cells also have cell walls. While the chief component of bacterial cell walls is peptidoglycan, the major organic molecule in the plant cell wall is cellulose (see structure below), a polysaccharide made up of glucose subunits.

Figure 9. Cellulose is a long chain of β-glucose molecules connected by a 1-4 linkage. The dashed lines at each end of the figure indicate a series of many more glucose units. The size of the page makes it impossible to portray an entire cellulose molecule.
Chloroplasts
Chloroplasts are plant cell organelles that carry out photosynthesis. Like the mitochondria, chloroplasts have their own DNA and ribosomes, but chloroplasts have an entirely different function.
Like mitochondria, chloroplasts have outer and inner membranes, but within the space enclosed by a chloroplast’s inner membrane is a set of interconnected and stacked fluid-filled membrane sacs called thylakoids (figure below). Each stack of thylakoids is called a granum (plural = grana). The fluid enclosed by the inner membrane that surrounds the grana is called the stroma.

Figure 10. The chloroplast has an outer membrane, an inner membrane, and membrane structures called thylakoids that are stacked into grana. The space inside the thylakoid membranes is called the thylakoid space. The light harvesting reactions take place in the thylakoid membranes, and the synthesis of sugar takes place in the fluid inside the inner membrane, which is called the stroma. Chloroplasts also have their own genome, which is contained on a single circular chromosome.
The chloroplasts contain a green pigment called chlorophyll, which captures the light energy that drives the reactions of photosynthesis. Like plant cells, photosynthetic protists also have chloroplasts. Some bacteria perform photosynthesis, but their chlorophyll is not relegated to an organelle.
Evolution connection
Endosymbiosis
We have mentioned that both mitochondria and chloroplasts contain DNA and ribosomes. Have you wondered why? Strong evidence points to endosymbiosis as the explanation.
Symbiosis is a relationship in which organisms from two separate species depend on each other for their survival. Endosymbiosis (endo- = “within”) is a mutually beneficial relationship in which one organism lives inside the other. Endosymbiotic relationships abound in nature. For instance, some microbes that live in our digestive tracks produce vitamin K. The relationship between these microbes and us (their hosts) is said to be mutually beneficial or symbiotic. The relationship is beneficial for us because we are unable to synthesize vitamin K; the microbes do it for us instead. The relationship is also beneficial for the microbes because they receive abundant food from the environment of the large intestine, and they are protected both from other organisms and from drying out.
Scientists have long noticed that bacteria, mitochondria, and chloroplasts are similar in size. We also know that bacteria have DNA and ribosomes, just as mitochondria and chloroplasts do. Scientists believe that host cells and bacteria formed an endosymbiotic relationship when the host cells ingested both aerobic and autotrophic bacteria (cyanobacteria) but did not destroy them. Through many millions of years of evolution, these ingested bacteria became more specialized in their functions, with the aerobic bacteria becoming mitochondria and the autotrophic bacteria becoming chloroplasts. There will be more on this later in the reading.
The central vacuole
Previously, we mentioned vacuoles as essential components of plant cells. If you look at the cartoon figure of the plant cell, you will see that it depicts a large central vacuole that occupies most of the area of the cell. The central vacuole plays a key role in regulating the cell’s concentration of water in changing environmental conditions.
Silly vacuole factoid: Have you ever noticed that if you forget to water a plant for a few days, it wilts? That’s because as the water concentration in the soil becomes lower than the water concentration in the plant, water moves out of the central vacuoles and cytoplasm. As the central vacuole shrinks, it leaves the cell wall unsupported. This loss of support to the cell walls of plant cells results in the wilted appearance of the plant.
The central vacuole also supports the expansion of the cell. When the central vacuole holds more water, the cell gets larger without having to invest a lot of energy in synthesizing new cytoplasm.
The scientific method overview
An example of oversimplification that confounds many students of biology (particularly early in their studies) is the use of language that hides the experimental process used to build knowledge. For the sake of expediency, we often tell stories about biological systems as if we are presenting unquestionable facts. However, while we often write and speak about topics in biology with a conviction that gives the appearance of "factual" knowledge, reality is often more nuanced and filled with significant uncertainties. The "factual" presentation of material (usually lacking discussion of evidence or confidence in the evidence) plays to our natural tendency to feel good about "knowing" things, but it tends to create a false sense of security in the state of knowledge and does little to encourage the use of imagination or the development of critical thinking.
A better way to describe our knowledge about the natural world would be to explicitly qualify that what we know to be "true" in science represents only our current best understanding of a topic; an understanding that has not yet been refuted by experiment. Unfortunately, repeated qualification becomes rather cumbersome. The important thing to remember is that while we may not say so explicitly, all of the knowledge we discuss in class represents only the best of our current understanding. Some ideas have withstood repeated and varied experimentation while other topics have yet to be tested as thoroughly. So if we're not as certain about things as we'd like to believe sometimes, how do we know what to put confidence in and what to be skeptical of? The complete answer is non-trivial but it begins with developing an understanding of the process we use in science to build new knowledge. The scientific method is the process by which new knowledge is developed. While the process can be described with long lists of "steps" (often seen in textbooks), its core elements can be described more succinctly.
Succinct description of scientific method (adapted from Feynman)
- Make an observation about the world.
- Propose a possible explanation for the observation.
- Test the explanation by experiment.
- If the explanation disagrees with experiment, the explanation is wrong.
At its core, that's it! In science there may be multiple, simultaneously proposed explanations or ideas that are tested by experiment. The ideas that fail experimentation are left behind. The ideas that survive experimentation move forward and are often retested by alternative experiments until they too either fail or continue to be retained.
Making an observation and asking a question
The ability to make useful observations and/or ask meaningful questions requires curiosity, creativity, and imagination—this cannot be overstated. Indeed, historically, it is first and foremost the application of these skills, perhaps more than technical ability, which has led to big advances in science. Many people think that making meaningful observations and asking useful questions is the easiest part of the scientific method. This is not always the case. Why? Seeing what others have not yet asked and creativity take work and thoughtful reflection! In addition, our senses of observation are often biased by life experience, prior knowledge, or even our own biology. These underlying biases influence how we see the world, how we interpret what we see, and what we are ultimately curious about. This means that when we look at the world, we can miss a lot of things that are actually right under our noses. Douglas Adams, who is best known for his book entitled The Hitchhiker’s Guide to the Galaxy, once expanded on this point by writing:
“The most misleading assumptions are the ones you don't even know you're making.”
Scientists, therefore, need to be aware of any underlying biases and any assumptions that may influence how they internalize and interpret observations. This includes approaching our bias that the variety of places we get our knowledge (i.e., textbooks, instructors, the Internet) are representing the absolute truth with a healthy dose of skepticism. We need to learn to examine the evidence underling the “facts” we supposedly know and make critical judgments about how much we trust that knowledge. More generally, taking the time to make careful observations and to uncover any assumptions and biases that could influence how they are interpreted is, therefore, time well spent. This skill, like all others, needs to be developed and takes practice and we’ll try to start you on this in BIS2A.
For fun, and to test your observation skills, Google “observation tests”. Many of the search results will take you to interesting psychological tests and/or videos that illustrate how difficult accurate observation can be.
Generating a testable hypothesis
The "possible explanation" referred to in step three above has a formal name; it is called a hypothesis. A hypothesis is not a random guess. A hypothesis is an educated (based on prior knowledge or a new viewpoint) explanation for an event or observation. It is typically most useful if a scientific hypothesis can be tested. This requires that the tools to make informative measurements on the system exist and that the experimenter has sufficient control over the system in question to make the necessary observations.
Most of the time, behaviors of the system that the experimenter wants to test can be influenced by many factors. We call the behaviors and factors dependent and independent variables, respectively. The dependent variable is the behavior that needs explaining while the independent variables are all of the other things that can change and influence the behavior of the dependent variable. For example, an experimenter that has developed a new drug to control blood pressure may want to test whether her new drug actually influences blood pressure. In this example, the system is the human body, the dependent variable might be blood pressure, and the independent variables might be other factors that change and influence blood pressure like age, sex, and levels of various soluble factors in the blood stream.
Note: for more on dependent and independent variables
Note
In BIS2A, and beyond, we prefer to avoid using language like “the experiment proved her hypothesis” when referring to a case like the blood pressure example above. Rather we would say, “the experiment is consistent with her hypothesis.” Note that for convenience, we referred to the alternative hypothesis simply as “her hypothesis”! It would be more correct to state, “the experiment falsified her null hypothesis and is consistent with her alternative hypothesis.” Why take this shortcut since doing so adds confusion when a student is trying to learn? In this case, it was done to illustrate the point above about language shortcuts and hence the lengthy explanation. However, be aware of this commonly used shortcut and learn to make sure you can read in the correct meaning yourself.
Note: possible discussion
What does the statement about falsifying hypotheses mean in your own words? Why is falsification critical to the scientific method?
Controls
In an ideal case, an experiment will include control groups. Control groups are experimental conditions in which the values of the independent variables (there may be more than one) are maintained as close to those in the experimental group with the exception of the independent variable being tested. In the blood pressure example, an ideal scenario would be to have one identical group of people taking the drug and another group of people identical to those in the experimental group taking a pill containing something known to not influence blood pressure. In this oversimplified example, all independent variables are identical in the control and experimental groups with the exception of the presence or absence of the new drug. Under these circumstances, if the value of the dependent variable (blood pressure) of the experimental group differs from that of the control group, one can reasonably conclude that the difference must be due to the difference in independent variable (the presence/absence of the drug). This is, of course, the ideal. In real life it is impossible to conduct the proposed drug dosage experiment; the sheer number of possible independent variables in a group of potential patients would be high. Fortunately, while statisticians have come to the rescue in real life, you won’t need to understand the nuances of these statistical issues in BIS2A.
Accuracy in measurement, uncertainty, and replication
Finally, we mention the intuitive notion that the tools used to make the measurements in an experiment must be reasonably accurate. How accurate? They must be accurate enough to make measurements with sufficient certainty to draw conclusions about whether changes in independent variables actually influence the value of a dependent variable. If we take, yet again, the blood pressure example above. In that experiment, we made the important assumption that the experimenter had tools that allowed her to make accurate measurements of the changes in blood pressure associated with the effects of the drug. For instance if the changes associated with the drug ranged between 0 and 3 mmHg and her meter capably measured changes in blood pressure with a certainty of +/- 5 mmHg, she could not have made the necessary measurements to test her hypothesis or would have missed seeing the effect of the drug. For the sake of the example, we assume that she had a better instrument and that she could be confident that any changes she measured were indeed differences due to the drug treatment and that they were not due to measurement error, sample-to-sample variability, or other sources of variation that lower the confidence of the conclusions that are drawn from the experiment.
The topic of measurement error leads us to mention that there are numerous other possible sources of uncertainty in experimental data that you as students will ultimately need to learn about. These sources of error have a lot to do with determining how certain we are that experiments have disproven a hypothesis, how much we should trust the interpretation of the experimental results and, by extension, our current state of knowledge. Even at this stage, you will recognize some experimental strategies used to deal with these sources of uncertainty (i.e., making measurements on multiple samples, creating replicate experiments). You will learn more about this in your statistics courses later on.
For now, you should, however, be aware that experiments carry a certain degree of confidence in the results and that the degree of confidence in the results can be influenced by many factors. Developing healthy skepticism involves, among other things, learning to assess the quality of an experiment and the interpretation of the findings and learning to ask questions about things like this.
Note: possible discussion
After moving to California to attend UC Davis, you have fallen in love with fresh tomatoes. You decide that the tomatoes in the stores just don’t taste right and resolve to grow your own.
You plant tomato plants all over your back yard; every free space now has a freshly planted tomato seedling of the same variety. You have planted tomatoes in the ground in full sunlight and next to your house in full shade.
Observation: After the first year of harvest, you make the observation that the plants growing in full shade almost always seem shorter than those in the full sun. You think that you have a reasonable explanation (hypothesis) for this observation.
Based on the information above, you create the following hypothesis to explain the differences in height you noticed in your tomatoes:
Hypothesis: The height that my tomato plants reach is positively correlated to the amount of sunlight they are exposed to (e.g., the more sun the plant gets, the taller it will be).
This hypothesis is testable and falsifiable. So, the next summer you decide to test your hypothesis.
This hypothesis also allows you to make a prediction. In this case you might predict that IF you were to shade a set of tomatoes in the sunny part of the yard, THEN those plants would be shorter than their full-sun neighbors.
You design an experiment to test your hypothesis by buying the same variety of tomato that you planted the previous year and plant your whole yard again. This year, however, you decide to do two different things:
- You create a shade structure that you place over a small subset of plants in the sunny part of your yard.
- You build a contraption with mirrors that redirects some sunlight onto a small subset of plants that are in the shady part of the yard.
Question 1: We used a shortcut above. Can you create statements for both the null and alternative hypothesis? Work with your classmates to do this.
Question 2: Why do you create a shade structure? What is this testing? Based on your hypothesis what do you predict will happen to the plants under the shade structure?
Question 3: Why do you create the mirror contraption? Why do you potentially need this contraption if you already have the shade structure?
New data: At the end of the summer you measure the height of your tomato plants and you find, once again, that the plants in the sunny part of the yard are indeed taller than those in the shady part of the yard. However, you notice that there is no difference in height between the plants under your shade structure and those right next to the structure in full sun. In addition, you notice that the plants in the shady part of the yard are all about the same height, including those that had extra light shined on them via your mirror contraption.
Question 4: What does this experiment lead you to conclude? What would you try to do next?
Question 5: Imagine an alternative scenario in which you discovered, as before, that the plants in the sunny part of the yard were all the same height (even those under your shade structure) but that the plants in the shady part of the yard that got “extra” light from your mirror contraption grew taller than their immediate neighbors. What would this say about your alternate hypothesis? Null hypothesis? What would you do next?
Question 6: What assumptions are you making about the ability to make measurements in this experiment? What influence might these assumptions have on your interpretation of the results?
In this class, you will occasionally be asked to create a hypotheses, to interpret data, and to design experiments with proper controls. All of these skills take practice to master—we can start to practice them in BIS2A. Again, while we don’t expect you to be masters after reading this text, we will assume that you have read this text during the first week and that the associated concepts are not completely new to you. You can always return to this text as a resource to refresh yourself.
Disclaimer
While the preceding treatment of the experimental method is very basic—you will undoubtedly add numerous layer of sophistication to these basic ideas as you continue in your studies—it should serve as a sufficient introduction to the topic for BIS2A. The most important point to remember from this section is that the knowledge represented in this course, while sometimes inadvertently represented as irrefutable fact, is really just the most current hypothesis about how certain things happen in biology that has yet to be falsified via experiment.
The Design Challenge
Your BIS2A instructors have devised something that we call “The Design Challenge” to help us approach the topics we cover in the course from a problem solving and/or design perspective. This teaching tool helps us:
• develop a frame of mind or way of approaching the material and
• design a set of sequential steps that help structure thinking about course topics in a problem-solving context.
How is it intended to work? When we encounter a topic in class, “The Design Challenge” encourages us to think about it in the following problem-solving centric way:
- Identify the problem(s) - this may include identifying "big" problems and also decomposing them into "smaller" nested sub-problems
- Determine criteria for successful solutions
- Identify and/or imagine possible solutions
- Evaluate the proposed solutions against the criteria for success
- Choose a solution
By using the structure of the design challenge, topics that are typically presented as lists of facts and stories are transformed into puzzles or problems that need solving. For instance the discussion about the topic of cell division is motivated by a problem. The problem statement can be: "The cell needs to divide". Some of the criteria for success can include needing to have a near identical copy of DNA in each daughter cell, distributing organelles between the daughter cells so that each remains viable etc. These would be considered sub-problems to the larger “the cell needs to divide” problem. One can then go on to explore what the challenges are and try to use existing knowledge and imagination to propose some solutions for each of those problems. Different solutions can be evaluated and then compared to what Nature seems to have done (at least in the cases that are well studied).
This exercise requires us to use imagination and critical thinking. It also encourages the student and instructor to think critically about why the particular topic is important to study. The Design Challenge approach to teaching biology attempts to make the student and instructor focus on the important core questions that drove the development of the knowledge in the first place! It also encourages students to dream up new ideas and to interact with the material in a manner that is question/problem-centered rather than “fact”-centered. The question/problem-centered approach is different from what most people are used to, but it is ultimately more useful for developing skills, mental frameworks and knowledge that will transfer to other problems that they will encounter during their studies and beyond.
Example
For example, the guiding problem in BIS2A is to understand “How to Build a Cell”. This rather complex problem will be broken down into several smaller sub-problems that include:
- acquiring the building blocks to construct cellular parts from the environment
- acquiring the energy to build cellular parts from the environment
- transforming the building blocks of the cell between different forms
- transferring energy between different storage forms
- creating a new cell from an old cell
- problems we identify in class
As we explore these sub-problems, we will at times explore some of the different ways in which biology has addressed each issue. As we get into details, let us however make sure to stay focused on and not forget the importance of always staying linked to the questions/problems that motivated us to talk about the specifics in the first plac
Scientific Method vs. The Design Challenge
At this point you might be thinking: "What is the difference between the scientific method and the design challenge rubric and why do I need both?" It's not an uncommon question so let's see if we can clarify this now.
The design challenge and the scientific method are both processes that share similar qualities. The critical distinguishing feature, however, is the purpose behind each of the processes. The scientific method is a process used for eliminating possible answers to questions. A typical scenario where one might use the scientific method would involve someone making an observation, proposing multiple explanations, designing an experiment that might help eliminate one or more of the explanations, and reflecting on the result. By contrast the design process is used for creating solutions to problems. A typical scenario for the design challenge would start with a problem that needs solving, defining criteria for a successful resolution, devising multiple possible solutions that would meet the success criteria and either selecting a solution or reflecting on changes that might be made to the designs to meet success criteria. A key operational difference is that the design challenge requires that criteria for success be defined while the scientific method does not.
While both are similar, the differences are still real and we need to practice both processes. We'll assert that we use both processes in "real life" all of the time. A physician, for instance, will use both processes interactively as she forms hypotheses that try to determine what might be causing her patient's ailments. She will turn around and use the design process to build a course of treatment that meets certain success criteria. A scientist may be deep into hypothesis generation but he will eventually need to use a design process for building an experiment that will, within certain definable success criteria, help him answer a question.
Both processes, while similar, are important to use in different situations and we want to begin getting better at both.
Models and simplifying assumptions
Creating models of real things
Life is complicated. To help us understand what we see around us—in both our everyday lives and in science or engineering—we often construct models. A common aphorism states: all models are wrong, but some are useful. That is, no matter how sophisticated, all models are approximations of something real. While they are not the “real thing” (and are thus wrong), models are useful when they allow us to make predictions about real life that we can use. Models come in a variety of forms that include, but are not limited to:
Types of models
- Physical models: These are 3-D objects that we can touch.
- Drawings: These can be on paper or on the computer and either in 2-D or virtual 3-D. We mostly look at them.
- Mathematical models: These describe something in real life in mathematical terms. We use these to calculate the behavior of the thing or process we want to understand.
- Verbal or written models: These models are communicated in written or spoken language.
- Mental models: These models are constructed in our minds and we use these to create the other types of models and to understand the things around us.
Simplifying assumptions
Usually, in science and everyday life alike, simple models are preferred over complex ones. Creating simple models of complex real things requires us to make what are known as simplifying assumptions. As their name implies, simplifying assumptions are assumptions that are included in the model to simplify the analysis as much as possible. When a simplified model no longer predicts behavior of the real thing within acceptable bounds, too many simplifying assumptions have been made. When little predictive value is gained from adding more details to a model, it is likely overly complex. Let’s take a look at different types of models from different disciplines and point out their simplifying assumptions.
An example from physics: a block on a frictionless plane

Figure 1. A line drawing that models a block (of any material) sitting on a generic incline plane. In this example some simplifying assumptions are made. For instance, the details of the materials of the block and plane are ignored. Often, we might also, for convenience, assume that the plane is frictionless. The simplifying assumptions allow the student to practice thinking about how to balance the forces acting on the block when it is elevated in a gravity field and to see that the surface it is sitting on is not perpendicular to the gravity vector (mg). This simplifies the math and allows the student to focus on the geometry of the model and how to represent that mathematically. The model, and its simplifying assumptions, might do a reasonably good job of predicting the behavior of an ice cube sliding down an glass incline plane but would likely do a bad job of predicting the behavior of a wet sponge on an incline plane coated with sand paper. The model would be oversimplified for the latter scenario.
Source: Created by Marc T. Facciotti (Own work)
An example from biology: a ribbon diagram of a protein—the transmembrane protein bacteriorhodopsin
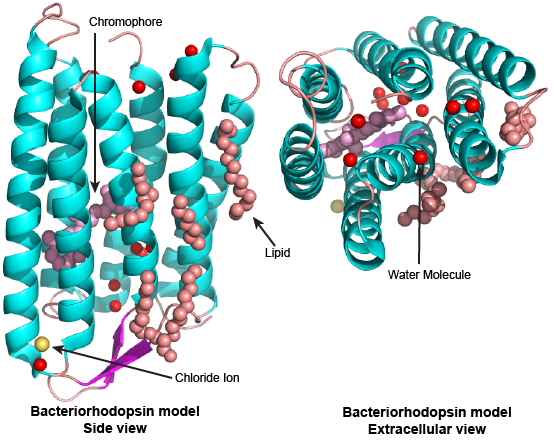
Figure 2. This is a cartoon model of the transmembrane protein bacteriorhodopsin. The protein is represented as a light blue and purple ribbon (the different colors highlight alpha helix and beta sheet, respectively), a chloride ion is represented as a yellow sphere, red spheres represent water molecules, pink balls and sticks represent a retinal molecule located on the "inside" of the protein, and orange balls-and-sticks represent other lipid molecules located on the "outside" surface of the protein. The model is displayed in two views. On the left the model is viewed "side on" while on the right it is viewed along its long axis from the extracellular side of the protein (rotated 90 deg out of the page from the view on the left). This model simplifies many of the atomic-level details of the protein. It also fails to represent the dynamics of the protein. The simplifying assumptions mean that the model would not do a good job predicting the time it takes for the protein to do its work or how many protons can be transported across a membrane per second. On the other hand, this model does a very good job of predicting how much space the protein will take up in a cellular membrane, how far into the membrane the retinal sits, or whether certain compounds can reasonably “leak” through the inner channel.
Source: Created by Marc T. Facciotti (own work), University of California, Davis
Derived from PDBID:4FPD
An example from chemistry: a molecular line model of glucose
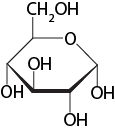
Figure 3. A line drawing of a glucose molecule. By convention, the points where straight lines meet are understood to represent carbon atoms while other atoms are shown explicitly. Given some additional information about the nature of the atoms that are figuratively represented here, this model can be useful for predicting some of the chemical properties of this molecule, including solubility or the potential reactions it might enter into with other molecules. The simplifying assumptions, however, hide the dynamics of the molecules.
Source: Created by Marc T. Facciotti (Own work)
An example from everyday life: a scale model of a Ferrari

Figure 4. A scale model of a Ferrari. There are many simplifications and most only make this useful for predicting the general shape and relative proportions of the real thing. For instance, this model gives us no predictive power about how well the car drives or how quickly it stops from a speed of 70 km/s.
Source: Created by Marc T. Facciotti (Own work)
Note: possible discussion
Describe a physical model that you use in everyday life. What does the model simplify from the real thing?
Note: possible discussion
Describe a drawing that you use in science class to model something real. What does the model simplify from the real thing? What are the advantages and disadvantages of the simplifications?
The spherical cow
The spherical cow is a famous metaphor in physics that make fun of physicists tendencies to create hugely simplified models for very complex things. Numerous jokes are associated with this metaphor and they go something like this:
"Milk production at a dairy farm was low, so the farmer wrote to the local university, asking for help from academia. A multidisciplinary team of professors was assembled, headed by a theoretical physicist, and two weeks of intensive on-site investigation took place. The scholars then returned to the university, notebooks crammed with data, where the task of writing the report was left to the team leader. Shortly thereafter the physicist returned to the farm, saying to the farmer, "I have the solution, but it only works in the case of spherical cows in a vacuum"."
Source: Wikipedia page on Spherical Cow - accessed November 23, 2015.

Figure 5. A cartoon representation of a spherical cow.
Source: https://upload.wikimedia.org/wikiped.../d2/Sphcow.jpg
By Ingrid Kallick (Own work) [GFDL (http://www.gnu.org/copyleft/fdl.html) or CC BY 3.0 (http://creativecommons.org/licenses/by/3.0)], via Wikimedia Commons
The spherical cow is an amusing way to ridicule the process of creating simple models and it is quite likely that you will have your BIS2A instructor invoke the reference to the spherical cow when an overly simplified model of something in biology is being discussed. Be ready for it!
Bounding or asymptotic analysis
In BIS2A, we use models frequently. Sometimes we also like to imagine or test how well our models actually represent reality and compare that with expectations from what we know to be true for the real life thing. There are many ways to do this depending on how precisely you need to know the behavior of the thing you're trying to model. If you need to know a lot of detail, you create a detailed model. If you're willing to live with less detail, you will create a simpler model. In addition to applying simplifying assumptions, it is often useful to assess your model using a technique we call bounding or asymptotic analysis. The main idea of this technique is to use the model, complete with simplifying assumptions, to understand how the real thing might behave at extreme conditions (e.g., evaluate the model at the minimum and maximum values of a variable). Let’s examine a simple real life example of how this technique works.
Example: bounding
Problem setup
Imagine that you need to leave Davis, CA and get home to Selma, CA for the weekend. It's 5PM and you told your parents that you'd be home by 6:30. Selma is 200 miles (322 kM) from Davis. You're getting worried that you won't make it home on time. Can you get some estimate of whether it's even possible or if you'll be reheating your dinner in the microwave?
Create simplified model and use of bounding
You can create a simplified model. In this case you can assume that the road between Davis and Selma is perfectly straight. You also assume that your car has only two speeds: 0 mph and 120 mph. These two speeds are the minimum and maximum speeds that you can travel—the bounding values. You can now estimate that even under assumptions of the theoretically "best case" scenario, where you would drive on a perfectly straight road with no obstacles or traffic at maximum speed, you will not make it home on time. At maximum speed you would only cover 180 of the required 200 miles in the 1.5 hours you have.
Interpretation
In this real life example a simplified model is created. In this case, one very important simplifying assumptions is made: the road is assumed to be straight and free of obstacles or traffic. These assumptions allow you to reasonably assume that you could drive this road at full speed the whole distance. The simplifying assumptions simplified out a lot of what you know is actually there in the real world that would influence the speed you could travel and by extension the time it would take to make the trip. The use of bounding—or calculating the behavior of at the minimum and maximum speeds—is a way of making quick predictions about what might happen in the real world.
We will conduct similar analyses in BIS2A.
The importance of knowing key model assumptions
Knowing what simplifying assumptions are made in a model is critical to judging how useful it is for predicting real life and for starting to make a guess about where the model needs improving if it is not sufficiently predictive. In BIS2A you will periodically be asked to create different types of models and to explicitly identify the simplifying assumptions and the impact of those assumptions on the utility and predictive ability of the model. We will also use models together with bounding exercises to try learning something about the potential behavior of a system.