
Enzymes
- Page ID
- 21285

Enzymes Section Overview
Enzymes are biological catalysts that speed up chemical reactions by lowering the activation energy. Enzymes are proteins comprising one or more polypeptide chains. Enzymes have an active site that provides a unique chemical environment made up of certain amino acid R groups (residues).
Enzyme action must
Enzymes
A substance that helps a chemical reaction to occur is a catalyst, and the special molecules that
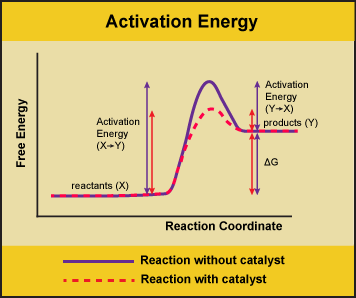
Figure 1. Enzymes lower the activation energy of the reaction but do not change the free energy of the reaction. Here, the solid line in the graph shows the energy required for reactants to turn into products without a catalyst. The dotted line shows the energy required using a catalyst. This figure should say Gibbs Free Energy on the Y-axis and instead of noting
Enzyme active site and substrate specificity
The chemical reactants to which an enzyme binds are the enzyme’s substrates. There may be one or more substrates, depending on the particular chemical reaction. In some reactions, a
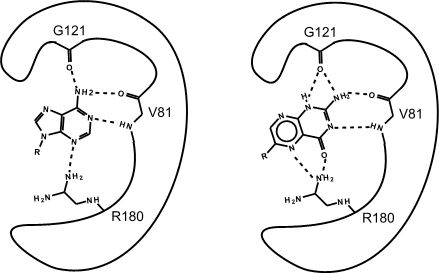
Figure 2. This is an enzyme with two different substrates bound in the active site.
At this point in the class, you should be familiar with all the
Possible NB Discussion Point: How your body breaks down caffeine
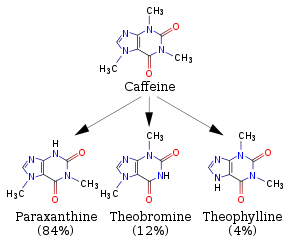
When you drink coffee or other caffeinated beverages like some sodas, you are consuming a molecule called caffeine! Caffeine over time gets metabolized (broken down) via a set of very related "CYP (Cytochrome P450)" enzymes to yield the three products shown in the figure below (Source: Wikipedia). To simplify slightly, you can interpret one arrow to represent a reaction catalyzed by one of the related CYP enzymes to yield paraxanthine, theobromine, or theophylline... all of which themselves get recognized by other enzymes that will further break them down and so on and so forth. Take a moment to examine the four structures below; the general structure should look vaguely familiar to you. Compare the reactant and the three products -- what are the noteworthy functional groups and properties of these molecules? What do you predict to be the key features of the active sites for the enzymes that break down these four molecules? If you were to design an enzyme that would break down caffeine AND theophylline only, how would you design your active site?
Exercise
Look to see which atoms in Figure 2 (
If you changed the pH of the solution that
Which substrate (the left or right one) do you think is more stable in the active site? Why? How?
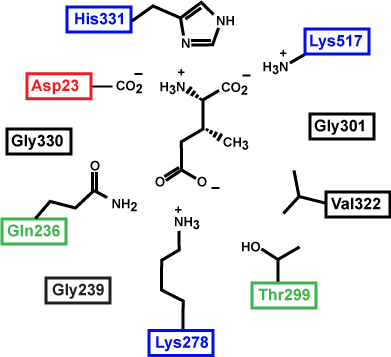
Figure 3. This is a depiction of an enzyme active site.
Source: created by
Exercise
First, identify the
A new way of visualizing diseases in the body.
Structural instability of enzymes
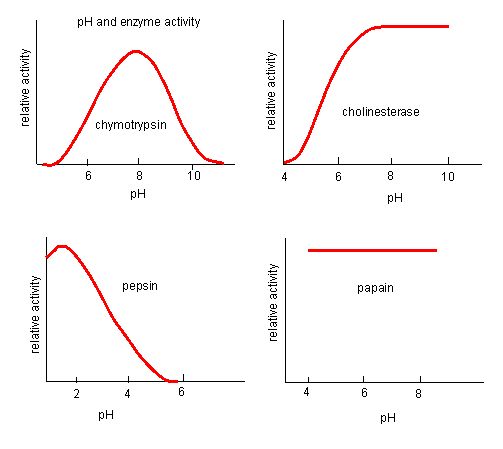
Figure 4. Enzymes have an optimal pH. The pH at which the enzyme is most active will be the pH where the active site R groups are
Source: http://biowiki.ucdavis.edu/Biochemis..._
The process where enzymes denature usually starts with the unwinding of the tertiary structure through destabilization of the bonds holding the tertiary structure together.
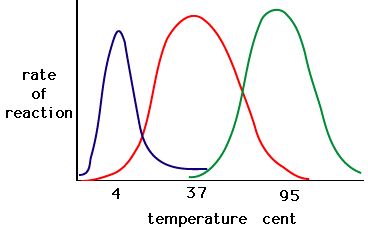
Figure 5. Enzymes have an optimal temperature. The temperature at which the enzyme is most active will usually be the temperature where the structure of the enzyme is stable or uncompromised. Some enzymes require a specific temperature to remain active and not denature. Source: http://academic.brooklyn.cuny.edu/bi..
Induced fit and enzyme function
For many years, scientists thought
When an enzyme binds its substrate, an enzyme-substrate complex
The activation energy required for many reactions includes the energy involved in slightly contorting chemical bonds so
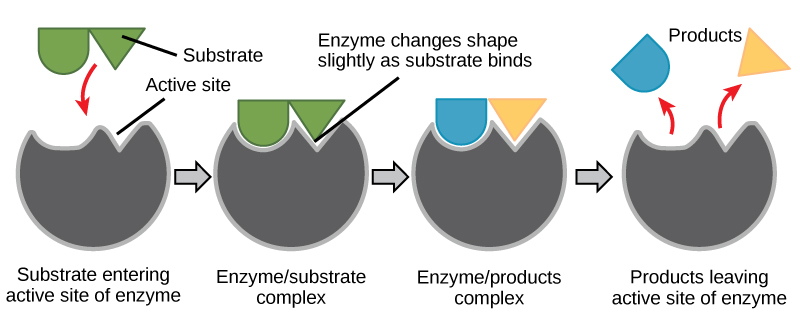
Figure 6. According to the induced-fit model, both enzyme and substrate undergo dynamic conformational changes upon binding. The enzyme contorts the substrate into its transition state,
Creating an energy story for the reaction above
Using Figure 6, answer the questions posed in the energy story.
1. What are the reactants? What are the products?
2. What work
3. What state is the energy in initially? What state is the energy transformed into in the final state? This one might be tricky still, but try to identify where the energy is in the initial state and the final state.
Enzyme regulation
Why regulate enzymes?
Cellular needs and conditions vary from cell to cell and change within individual cells over time. The required enzymes and energetic demands of stomach cells are different from those of fat storage cells, skin cells, blood cells, and nerve cells. Furthermore, a digestive cell works much harder to process and break down nutrients during the time closely following a meal compared with many hours after a meal. As these cellular demands and conditions vary, so do the needed amounts and functionality of different enzymes.
Regulation of enzymes by molecules
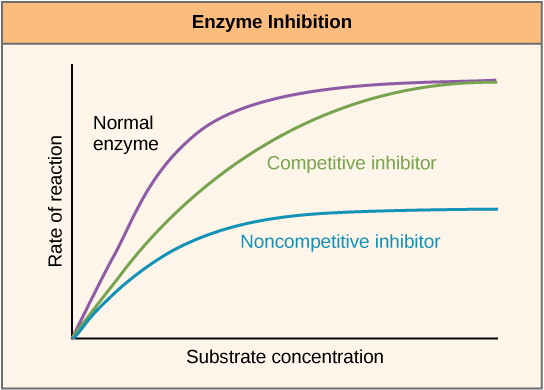
Figure 7. Competitive and noncompetitive inhibition affect the rate of reaction differently. Competitive inhibitors affect the initial rate but do not affect the maximal rate, whereas noncompetitive inhibitors affect the maximal rate.
Some inhibitor molecules bind to enzymes in a location where their binding induces a conformational change that reduces the affinity of the enzyme for its substrate. This
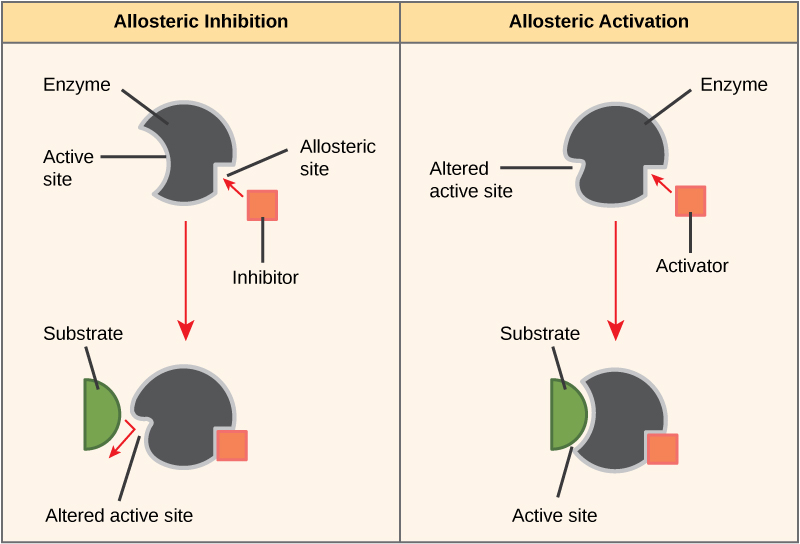
Figure 8. Allosteric inhibitors modify the active site of the enzyme so that substrate binding is reduced or prevented. In contrast, allosteric activators modify the active site of the enzyme so that the affinity for the substrate increases.
Video link
Check out this short (one-minute) video on competitive vs. noncompetitive enzymatic inhibition. Also, take a look at this video (1.2 minutes) on feedback inhibition.
Many enzymes don’t work optimally, or even at all, unless bound to other specific non-protein helper molecules, either temporarily through ionic or hydrogen bonds or permanently through stronger covalent bonds. Two types of helper molecules are cofactors and coenzymes. Binding to these molecules promotes optimal conformation and function for their respective enzymes. Cofactors are inorganic ions such as iron(II) (Fe2+) and magnesium(II) (Mg2+). One example of an enzyme that requires a metal ion as a cofactor is the enzyme that builds DNA molecules, DNA polymerase, which requires a bound zinc(II) ion (Zn2+) to function. Coenzymes are organic helper molecules, with a basic atomic structure made up of carbon and hydrogen, that are required for enzyme action. The most common sources of coenzymes are dietary vitamins. Some vitamins are precursors to coenzymes, and others act directly as coenzymes. Vitamin C is a coenzyme for multiple enzymes that take part in building the important connective tissue component, collagen. An important step in the breakdown of glucose to yield energy is catalysis by a multi-enzyme complex called pyruvate dehydrogenase. Pyruvate dehydrogenase is a complex of several enzymes that actually requires one cofactor (a magnesium ion) and five different organic coenzymes to catalyze its specific chemical reaction. Therefore, enzyme function is, in part, regulated by an abundance of various cofactors and coenzymes, which are supplied primarily by the diets of most organisms.
Enzyme compartmentalization
In eukaryotic cells, molecules such as enzymes are usually compartmentalized into different organelles. This allows for yet another level of regulation of enzyme activity. Enzymes required only for certain cellular processes can be housed separately along with their substrates, allowing for more efficient chemical reactions. Examples of this sort of enzyme regulation based on location and proximity include the enzymes involved in the latter stages of cellular respiration, which take place exclusively in the mitochondria, and the enzymes involved in the digestion of cellular debris and foreign materials, located within lysosomes.
Possible NB Discussion Point: Reversing the Effects of Caffeine
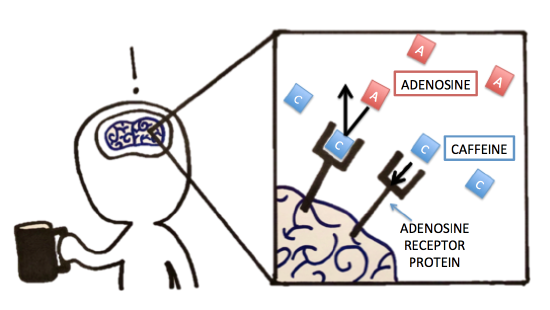
Previously, we discussed caffeine and its metabolism. Let’s now think about caffeine’s pharmacology (mode of action). Were you able to identify, compare, and contrast the molecule that caffeine had a similar structure to? Because of caffeine's structural similarity to the molecule adenosine, it is actually able to bind to the adenosine-specific receptor protein in the brain. However, because the exact lock-and-key-fit is unsatisfied, caffeine will not "activate" the adenosine receptors upon binding as adenosine would. Normally, when adenosine binds to and thereby activates its specific receptor protein in the brain, the physiological effect is increased drowsiness and muscle relaxation. It makes sense that we get tired at night because we accumulate adenosine over the day -- that's a lot of receptor activation! But back to caffeine -- when caffeine is present, it can bind to the adenosine receptor protein, thereby blocking adenosine from binding/activating the receptor. The lack of adenosine action is what leads to suppressed sleepiness and increased alertness. The inhibition seen with this receptor protein and caffeine is similar to some of the inhibition we see with enzymes. What type of inhibition would you classify this as? Follow up question: If you were hired by a company to design a solution to reverse the effect of caffeine post-ingestion, what strategies would you try to test? Explain!
Additional links
Khan Academy
The following links will take you to a series of videos on kinetics. The first link contains four videos on reaction rates, and the second link contains nine videos related to the relationship between reaction rates and concentration. These videos are supplemental and