
A Hypothesis for How ETC May Have Evolved*#
- Page ID
- 21254

A Hypothesis for How ETCs May Have Evolved
A proposed link between SLP/fermentation and the evolution of ETCs:
In a previous discussion of energy metabolism, we explored substrate level phosphorylation (SLP) and fermentation reactions. While SLP and fermentation together are perfectly good ways to harvest energy, one byproduct of these reactions is the acidification of the cell. Early cells that used these modes of energy harvested therefore needed to
In the scheme proposed above, one potential source of "wasted ATP" is its use in the removal of protons from the cell's cytosol; organisms that could find other mechanisms to expel accumulating protons while still preserving ATP could have a selective advantage. We hypothesize that this selective evolutionary pressure potentially led to the evolution of the first membrane-bound proteins that used red/ox reactions as their energy source (depicted in a second picture) to pump out the accumulating protons. Enzymes and enzyme complexes with these properties exist today in the as electron transport complexes like Complex I, the NADH dehydrogenase.
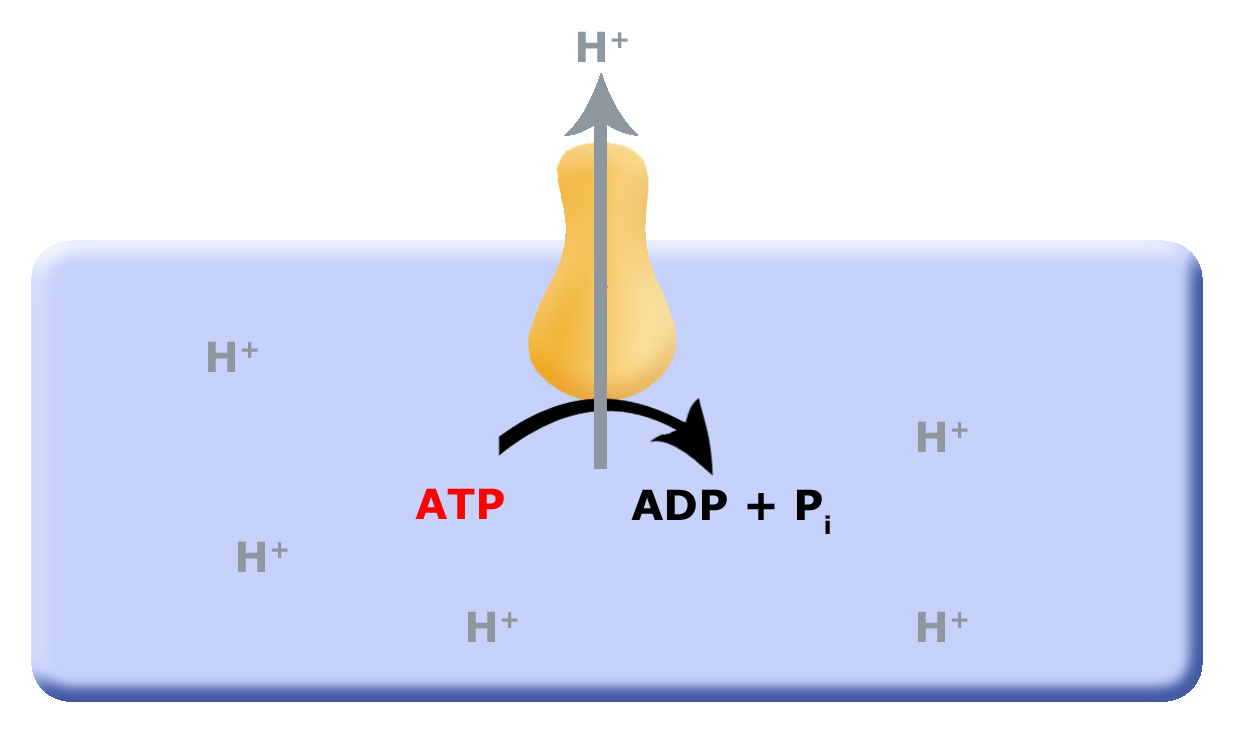
Figure 1. Proposed evolution of an
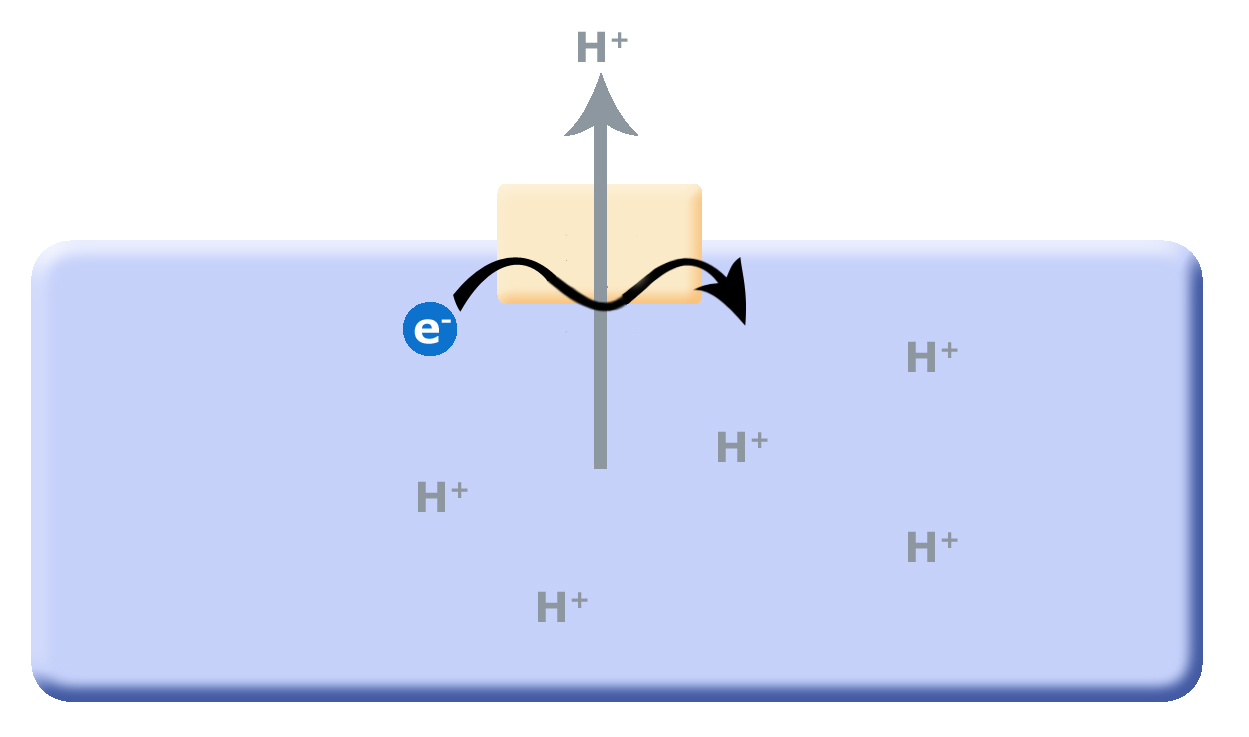
Figure 2. As small reduced organic molecules become limited, organisms that can find alternative mechanisms to remove protons from the cytosol may have
Continuing with this line of logic, if organisms evolved that could now use

Figure 3. The evolution of the
Note: Extended reading on the evolution of electron transport chains
If you're interested in
Possible NB Discussion  Point
Point
Dinitrophenol (DNP) is a small chemical that serves to uncouple the flow of protons across the inner mitochondrial membrane to the ATP synthase, making the membrane leaky to protons. People used it until 1938 as a weight-loss drug. What effect would you expect DNP to have on the difference in pH across both sides of the inner mitochondrial membrane? Why do you think this might be an effective weight-loss drug? Why might it be dangerous? Can you think of any scenarios where it is non-harmful, or even beneficial, to uncouple proton flow with ATP synthase?