
4.17: 2020_Spring_Bis2a_Facciotti_Lecture_17
- Page ID
- 27859
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)
Learning goals associated with 2020_Spring_Bis2A_Facciotti_Lecture_17
|
Light Energy and Pigments
Light Energy
The sun emits an enormous amount of electromagnetic radiation (solar energy) that spans a broad swath of the electromagnetic spectrum, the range of all radiation frequencies. When solar radiation reaches Earth, a fraction of this energy interacts with and may transfer to the matter on the planet. This energy transfer results in a wide variety of different phenomena, from influencing weather patterns to driving myriad biological processes. In BIS2A, we
First,
- Light in a vacuum travels at a constant speed of 299,792,458
m /s. We often abbreviate the speed of light with the variable "c ". - Light has properties of waves. A specific "color" of light has a characteristic wavelength.
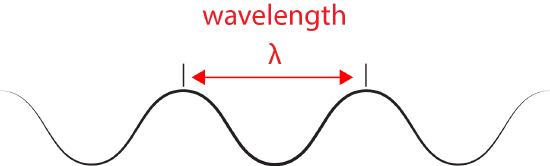
We refer to the distance between peaks in a wave as the wavelength and abbreviate it with the Greek letter lambda (Ⲗ). Attribution:
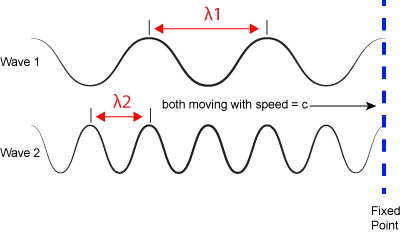
The inverse proportionality of frequency and wavelength. Wave 1 has a wavelength that is 2x that of wave 2 (Ⲗ1 > Ⲗ2). If the two waves are traveling at the same speed (
3. Finally, each frequency (or wavelength) of light
\
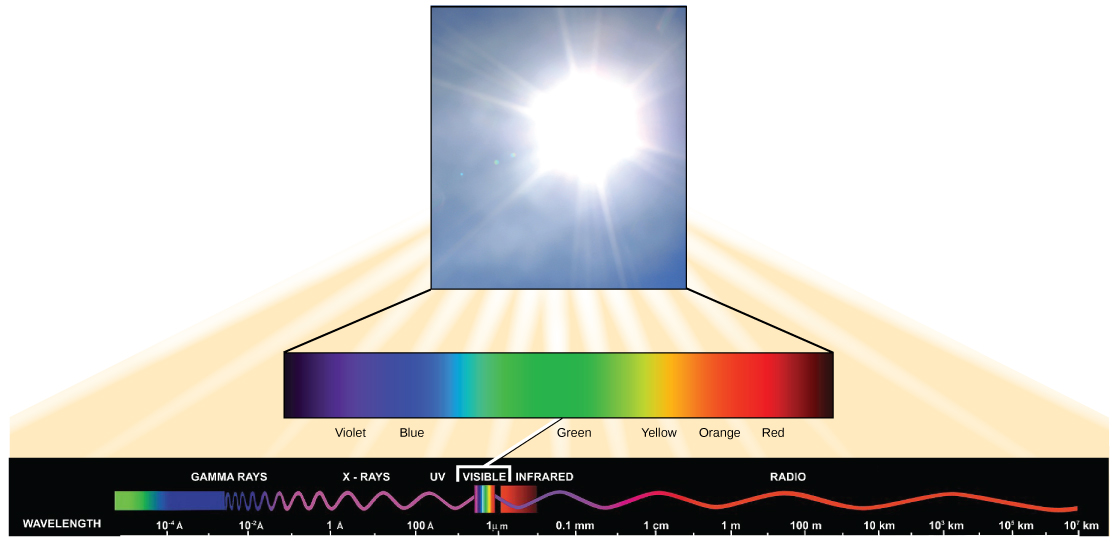
The sun emits energy in the form of electromagnetic radiation. All electromagnetic radiation, including visible light,
The Light We See
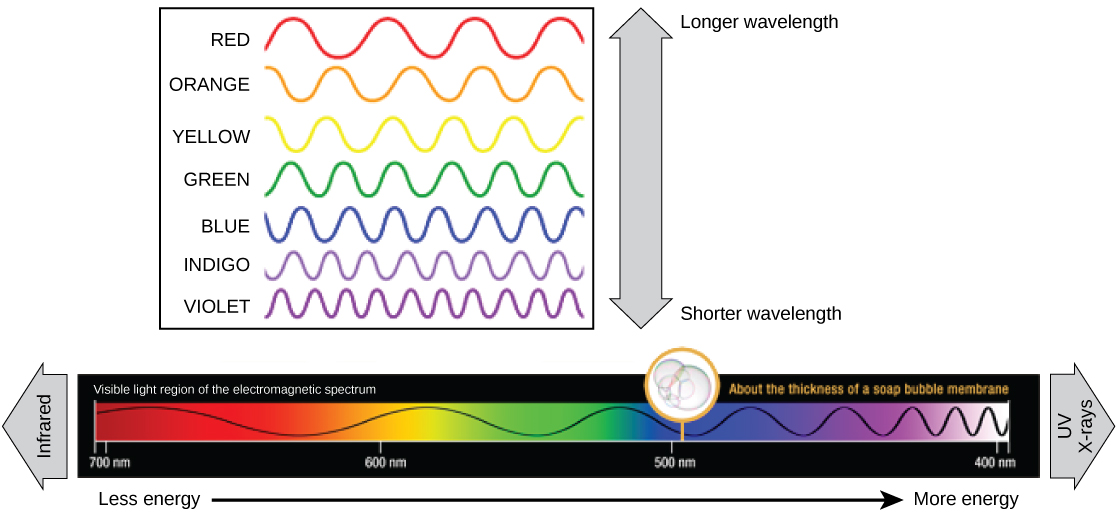
The colors of visible light do not carry the same amount of energy. Violet has the shortest wavelength and therefore carries the most energy, whereas red has the longest wavelength and carries the least amount of energy. Credit: modification of work by
Absorption by Pigments
The interaction between light and biological systems occurs through several mechanisms, some of which you may learn about in upper division courses in cellular physiology or biophysical chemistry. In BIS2A, we concern ourselves mostly with the interaction between light and biological pigments. These interactions can start a variety of light-dependent biological processes that can
Possible NB Discussion  Point
Point
Photophosphorylation is a process involving an electron transport chain that allows organisms to harvest energy from light. Some of you may already be familiar with this process. Many of you are learning about this for the first time. Given your current knowledge base, offer your best explanation or hypothesis as to how light interacts with the ETC. You will have a chance to revisit this topic very soon.
Three Key Types of Pigments We Discuss in BIS2A
Chlorophylls
Chlorophylls (including bacteriochlorophylls) are part of a large family of pigment molecules. There are five major chlorophyll pigments named:
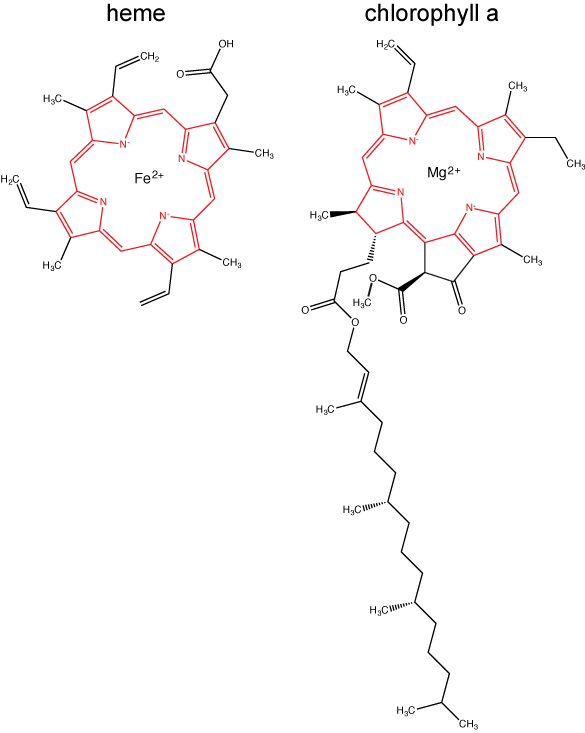
The structure of heme and chlorophyll
Carotenoids
Carotenoids are the red/orange/yellow pigments found in nature. They
Flavonoids
Flavonoids are a very broad class of compounds that
The core ring structure of
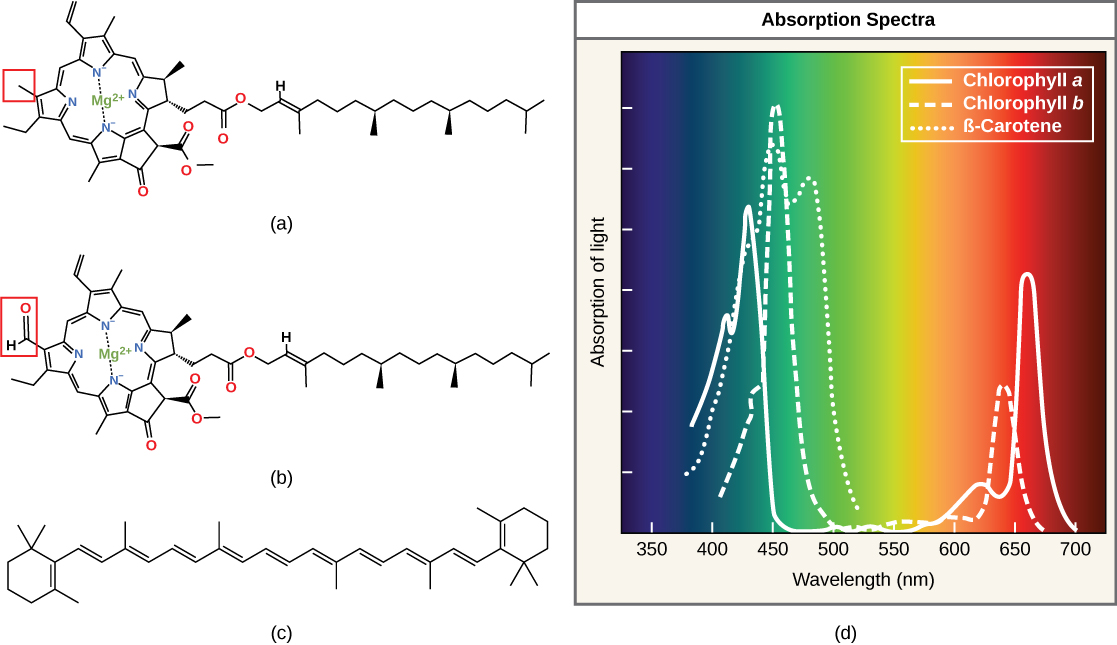
(a) Chlorophyll
Importance of having multiple different pigments
Not all photosynthetic organisms have full access to sunlight. Some organisms grow underwater where light intensity and the number of wavelengths decrease and change, respectively, with depth. Other organisms grow in competition for light. For instance, plants on the rainforest floor must be able to absorb any bit of light that comes through because the taller trees absorb most of the sunlight and scatter the remaining solar radiation. To account for these variable light conditions, many photosynthetic organisms have a mixture of pigments whose expression they can tune to improve the organism's ability to absorb energy from a wider range of wavelengths than would be possible with one pigment alone.
Photophosphorylation
Photophosphorylation an overview
Photophosphorylation is
As electrons pass from one electron carrier to another via red/ox reactions, enzymes can couple these exergonic electron transfers to the endergonic transport (or pumping) of protons across a membrane to create an electrochemical gradient. This electrochemical gradient generates a proton motive force (PMF). Enzymes can couple the exergonic drive of these protons to reach equilibrium to the endergonic production of ATP, via ATP synthase. As we will see in more detail, the electrons involved in this electron transport chain can have one of two fates: (1) they may return to their initial source in a process called cyclic photophosphorylation; or (2) they can transfer onto a close relative of NAD+ called NADP+. If electrons return to the original pigment in a cyclic process, the whole process can start over. If, however, the electron transfers onto NADP+ to form NADPH (**shortcut note—we didn't explicitly mention any protons but assume that they
What happens when a compound absorbs a photon of light?
When a compound absorbs a photon of light, the compound is said to leave its ground state and become "excited".

What are the fates of the "excited" electron? There are four
- The
e - can relax to a lower quantum state, transferring energy as heat. - The
e - can relax to a lower quantum state and transfer energy into a photon of light—a process known as fluorescence. The energy can be transferred by resonance to a neighboring molecule as thee - returns to a lower quantum state.- The energy can change the reduction potential such that the molecule can become an
e - donor. Linking this excitede - donor to a propere - acceptor can lead to an exergonic electron transfer.The excited state can be involved in red/ox reactions.

As the excited electron decays back to its lower energy state, it can transfer its energy in a variety of ways. While many so-called antenna or auxiliary pigments absorb light energy and transfer it to something known as a reaction center (by mechanisms depicted in option III in Figure 2), it is what happens at the reaction center that we are most concerned with (option IV in the figure above). Here a chlorophyll or bacteriochlorophyll molecule absorbs a photon's energy, and an electron is excited. This energy transfer suffices to allow the reaction center to donate the electron in a red/ox reaction to a second molecule. This starts the electron transport reactions. The result is an oxidized reaction center that must now
Simple photophosphorylation systems: anoxygenic photophosphorylation
Early in the evolution of photophosphorylation, these reactions evolved in anaerobic environments where there was very little molecular oxygen available. Two sets of reactions evolved under these conditions, both directly from anaerobic respiratory chains as described previously. We know these as the light reactions because they require the activation of an electron (an "excited" electron) from the absorption of a photon of light by a reaction center pigment, such as bacteriochlorophyll. We classify the light reactions either as cyclic or as noncyclic photophosphorylation, depending upon the final state of the electron
|
oxidized form |
|
|
Eo´ (volts) |
|---|---|---|---|
|
PS1* (ox) |
PS1* (red) |
- |
-1.20 |
|
ferredoxin (ox) version 1 |
ferredoxin (red) version 1 |
1 |
-0.7 |
|
PSII* (ox) |
PSII* (red) |
- |
-0.67 |
|
P840* (ox) |
PS840* (red) |
- |
-0.67 |
|
acetate |
acetaldehyde |
2 |
-0.6 |
|
CO2 |
Glucose |
24 |
-0.43 |
|
ferredoxin (ox) version 2 |
ferredoxin (red) version 2 |
1 |
-0.43 |
|
CO2 |
formate |
2 |
-0.42 |
|
2H+ |
H2 |
2 |
-0.42 (at [H+] = 10-7; |
|
NAD+ + 2H+ |
NADH + H+ |
2 |
-0.32 |
|
NADP+ + 2H+ |
NADPH + H+ |
2 |
-0.32 |
|
Complex I FMN (enzyme bound) |
FMNH2 |
2 |
-0.3 |
|
|
Lipoic acid, (red) |
2 |
-0.29 |
|
FAD+ (free) + 2H+ |
FADH2 |
2 |
-0.22 |
|
Pyruvate + 2H+ |
lactate |
2 |
-0.19 |
|
FAD+ + 2H+ (bound) |
FADH2 (bound) |
2 |
0.003-0.09 |
|
|
UQH. |
1 |
0.031 |
|
UQ + 2H+ |
UQH2 |
2 |
0.06 |
|
Plastoquinone; (ox) |
Plastoquinone; (red) |
- |
0.08 |
|
Ubiquinone; (ox) |
Ubiquinone; (red) |
2 |
0.1 |
|
Complex III Cytochrome b2; Fe3 |
Cytochrome b2; Fe2 |
1 |
0.12 |
|
Complex III Cytochrome c1; Fe3 |
Cytochrome c1; Fe2 |
1 |
0.22 |
|
Cytochrome |
Cytochrome c; Fe2 |
1 |
0.25 |
|
Complex IV Cytochrome |
Cytochrome |
1 |
0.29 |
|
1/2 O2 + H2O |
H2O2 |
2 |
0.3 |
|
P840GS (ox) |
PS840GS (red) |
- |
0.33 |
|
Complex IV Cytochrome a3; Fe3 |
Cytochrome a3; Fe2 |
1 |
0.35 |
|
Ferricyanide |
ferrocyanide |
2 |
0.36 |
|
Cytochrome |
Cytochrome f; Fe2 |
1 |
0.37 |
|
PSIGS (ox) |
PSIGS (red) |
. |
0.37 |
|
Nitrate |
nitrite |
1 |
0.42 |
|
Fe3+ |
Fe2+ |
1 |
0.77 |
|
1/2 O2 + 2H+ |
H2O |
2 |
0.816 |
|
PSIIGS (ox) |
PSIIGS (red) |
- |
1.10 |
|
* Excited State, after absorbing a photon of light GS Ground State, PS1: Oxygenic photosystem I P840: Bacterial reaction center containing bacteriochlorophyll (anoxygenic) PSII: Oxygenic photosystem II |
|||
Cyclic photophosphorylation
In cyclic photophosphorylation the bacteriochlorophyllred molecule absorbs enough light energy to energize and eject an electron to form bacteriochlorophyllox. The electron reduces a carrier molecule in the reaction center which

Figure 4. Cyclic electron flow. The reaction center P840 absorbs light energy and becomes excited, denoted with an *. The excited electron
Possible NB Discussion Point
The figure of cyclic photophosphorylation above depicts the flow of electrons in a respiratory chain. How does this process help generate ATP? Why might running the process in a cyclical fashion be advantageous for a cell?
Noncyclic photophosphorylation
In cyclic photophosphorylation, electrons cycle from bacteriochlorophyll (or chlorophyll) to a series of electron carriers and eventually back to bacteriochlorophyll (or chlorophyll); there is theoretically no net loss of electrons and they stay in the system. In noncyclic photophosphorylation, electrons leave from the photosystem and red/ox chain and eventually end up on NADPH. That means there needs to be a source of electrons, a source that has a smaller reduction potential than bacteriochlorophyll (or chlorophyll) that can donate electrons to bacteriochlorophyllox to reduce it. From looking at the electron tower in Figure 3, you can see what compounds can reduce the oxidized form of bacteriochlorophyll. The second requirement is that, when bacteriochlorophyll becomes oxidized by ejecting its excited electron, it must reduce a carrier that has a greater reduction potential than NADP/NADPH (see the electron tower). Here, electrons can flow from energized bacteriochlorophyll to NADP forming NADPH and oxidized bacteriochlorophyll. The system loses electrons and they end up on NADPH; to complete the circuit, bacteriochlorophyllox
Noncyclic electron flow

Figure 5. Noncyclic electron flow. In this example, the P840 reaction center absorbs light energy and becomes energized; the emitted electron reduces a FeS protein and reduces ferridoxin. Reduced ferridoxin (
We note that for bacterial photophosphorylation pathways, for each electron donated from a reaction center [remember only one electron is actually donated to the reaction center (or chlorophyl molecule)], the resulting output from that electron transport chain is either the formation of NADPH (requires two electrons) or ATP can be made but NOT not both. The path the electrons take in the ETC can have one or two outcomes. This puts limits on the versatility of the bacterial anoxygenic photosynthetic systems. But what would happen if a process evolved that used both systems? More precicely, a cyclic and noncyclic photosynthetic pathway which could form both ATP and NADPH from a single input of electrons? A second limitation is that these bacterial systems require compounds such as reduced sulfur to act as electron donors to reduce the oxidized reaction centers, but they are not necessarily widely found compounds. What would happen if a chlorophyll
Oxygenic photophosphorylation
Generation of NADPH and ATP
The overall function of light-dependent reactions is to transfer solar energy into chemical compounds, largely the molecules NADPH and ATP. This energy supports the light-independent reactions and fuels the assembly of sugar molecules. We depict the light-dependent reactions in Figures 6 and 7. Protein complexes and pigment molecules work together to produce NADPH and ATP.

The actual step that transfers light energy into a biomolecule takes place in a multiprotein complex called a photosystem, two types of which
Both photosystems have the same basic structure; several antenna proteins to which the chlorophyll molecules bind surround the reaction center in which the photochemistry takes place. Each photosystem associates with the light-harvesting complex, which passes energy captured from sunlight to the reaction center; it comprises multiple antenna proteins that contain a mixture of 300–400 chlorophyll
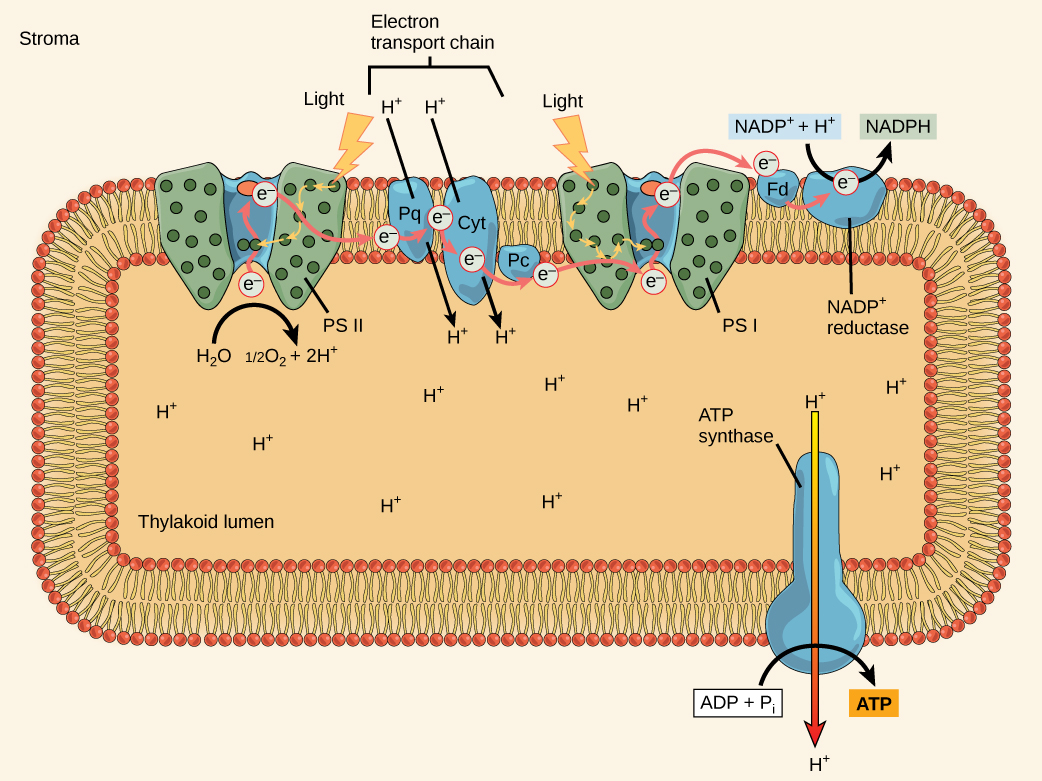
The reaction center contains a pair of chlorophyll
The Z-scheme
PSII and PSI are two major components of the photosynthetic electron transport chain, which also include the cytochrome complex. The reaction center of PSII (called P680) delivers its high-energy electrons, one at a time, to a primary electron acceptor called pheophytin (Ph), and then sequentially to two bound plastoquinones QA and QB. Electrons then transfer off of PSII onto a pool of mobile plastoquinones (Q pool) which then transfer the electrons to a protein complex called Cytochromeb6f. The cytochrome complex uses the red/ox transfers to pump proton across the thylakoyd membrane establishing a proton-motive force that can
As electrons move through the proteins that live between PSII and PSI, they take part in exergonic red/ox transfers. The free energy associated with the exergonic red/ox reaction

Figure 3. A diagram depicting the flow of electrons and the red/ox potentials of their carriers in oxygenic photosynthetic systems expressing both photosystem I (boxed in blue) and photosystem II (boxed in green). Ph = pheophytin; QA = bound plastoquinone, QB = more loosely associated plastoquinone; Q pool = mobile plastoquinone pool;
Light Independent Reactions and Carbon Fixation
A short introduction
The general principle of carbon fixation is that some cells under certain conditions can take inorganic carbon, CO2 (also referred to as mineralized carbon), and reduce it to a usable cellular form. Most of us know that green plants can take up CO2 and produce O2 in a process known as photosynthesis. We have already discussed photophosphorylation, the ability of a cell to transfer light energy onto chemicals and ultimately to produce the energy carriers ATP and NADPH in a process known as the light reactions. In photosynthesis, the plant cells use the ATP and NADPH formed during photophosphorylation to reduce CO2 to sugar, (as we will see, specifically G3P) in what we call the dark reactions. While we appreciate that this process happens in green plants, photosynthesis had its evolutionary origins in the bacterial world. In this module we will go over the general reactions of the Calvin Cycle, a reductive pathway that incorporates CO2 into cellular material.
In photosynthetic bacteria, such as Cyanobacteria and purple non-sulfur bacteria, as well plants, the energy (ATP) and reducing power (NADPH) - a term used to describe electron carriers in their reduced state -
The Calvin Cycle: the reduction of CO2 to Glyceraldehyde 3-Phosphate
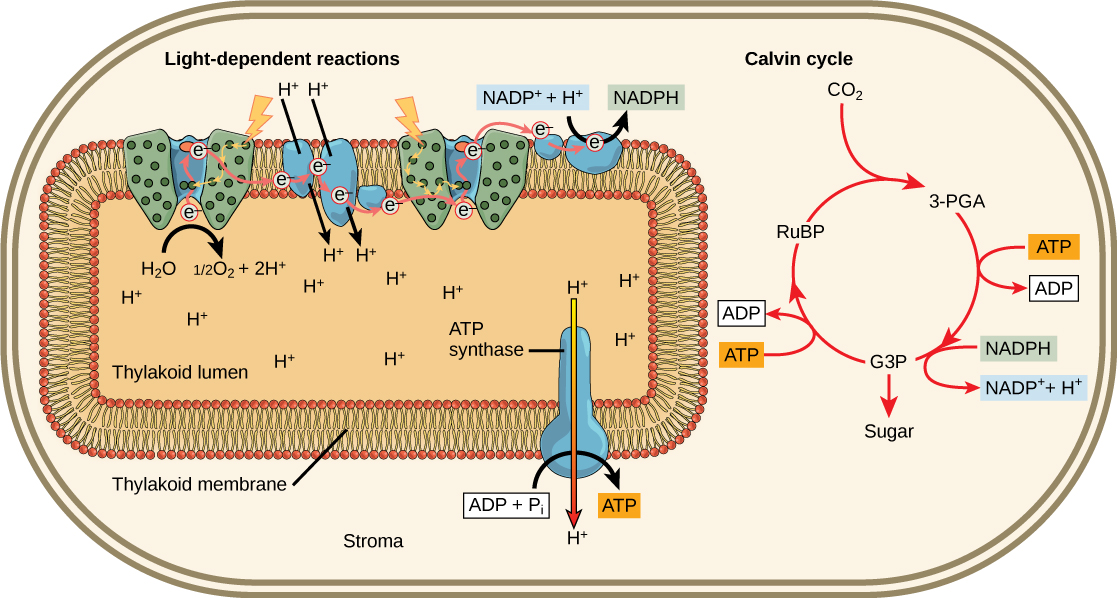
Figure 1. Light reactions harness energy from the sun to produce chemical bonds, ATP, and NADPH.
In plant cells,
Stage 1: Carbon Fixation
In the stroma of plant chloroplasts, besides CO2, two other components are present to start the light-independent reactions: an enzyme called ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO), and three molecules of ribulose bisphosphate (RuBP), as shown in the figure below. Ribulose-1,5-bisphosphate (RuBP) is composed of five carbon atoms and includes two phosphates.
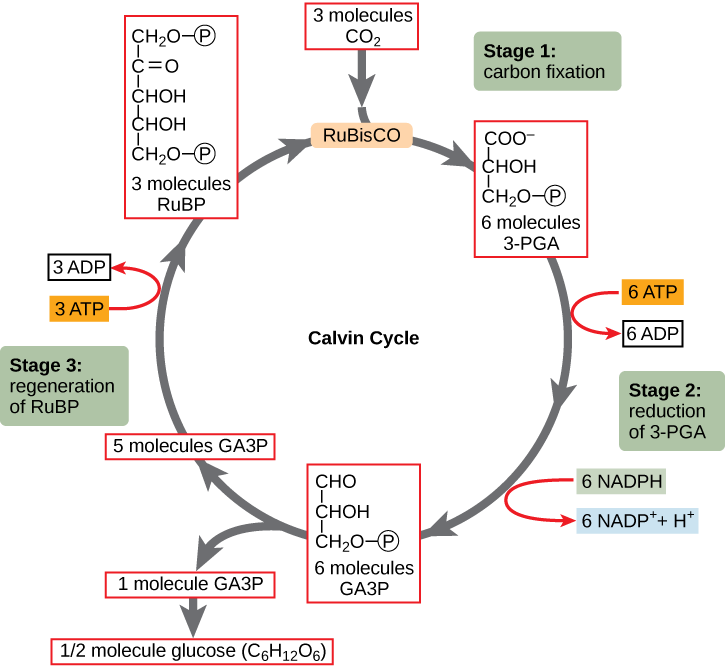
Stage 2: Reduction
ATP and NADPH are used to convert the six molecules of 3-PGA into six molecules of a chemical called glyceraldehyde 3-phosphate (G3P) - a carbon compound also found in glycolysis. The process uses six molecules of both ATP and NADPH. The exergonic process of ATP hydrolysis is in effect driving the endergonic redox reactions, creating ADP and NADP+. Both "spent" molecules (ADP and NADP+) return to the nearby light-dependent reactions to
Stage 3: Regeneration
Interestingly,
Possible NB Discussion Point
Have you ever heard anyone accidentally refer to the Amazon rainforest as the "lungs of the Earth"? In reality, the majority of our planet's oxygen is produced by marine organisms, such as microscopic phytoplankton -- which, by the way, also take up appreciable amounts of carbon dioxide from the environment. The family of phytoplankton include organisms like cyanobacteria and diatoms (a visually stunning type of algae -- look it up!) that are able to survive and aggregate close to the water's surface, where sun exposure is higher. Try to approach phytoplankton from a BIS 2A lens... What biochemical processes had to happen in order for these phytoplankton to produce oxygen? What exactly are the phytoplankton doing with the carbon dioxide they take up from the atmosphere? What large-scale global effects would you expect if phytoplankton health were to be severely compromised?