
3.5: DNA Repair
- Page ID
- 39841
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Learning Objectives
- Describe and compare direct repair, base excision repair, nucleotide excision repair, and mismatch repair pathways.
- Understand the situations in which non-homologous end-joining and homologous recombination are used for repair.
Maintaining the Integrity of the Cell's Information: DNA Repair
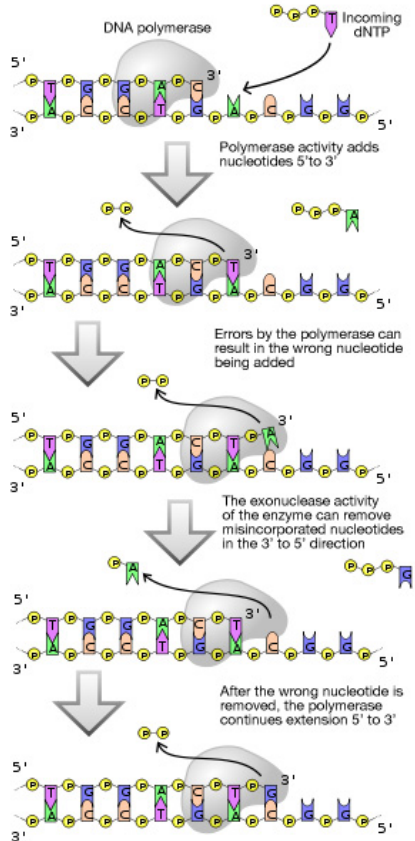
If DNA is the master copy of instructions for an organism, then it is important not to make mistakes when copying the DNA to pass on to new cells. Although proofreading by DNA polymerases greatly increases the accuracy of replication, there are additional mechanisms in cells to further ensure that newly replicated DNA is a faithful copy of the original, and also to repair damage to DNA during the normal life of a cell.
All DNA suffers damage over time, from exposure to ultraviolet and other radiation, as well as from various chemicals in the environment. Even chemical reactions naturally occurring within cells can give rise to compounds that can damage DNA. As you already know, even minor changes in DNA sequence, such as point mutations can sometimes have far-reaching consequences. Likewise, unrepaired damage caused by radiation, environmental chemicals or even normal cellular chemistry can interfere with the accurate transmission of information in DNA. Maintaining the integrity of the cell's "blueprint" is of vital importance and this is reflected in the numerous mechanisms that exist to repair mistakes and damage in DNA.
Post-Replicative Mismatch Repair
Does proofreading eliminate all errors made during replication? No. While proof-reading significantly reduces the error rate, not all mistakes are fixed on the fly by DNA polymerases. What mechanisms exist to correct the replication errors that are missed by the proof-reading function of DNA polymerases?
Errors that slip by proofreading during replication can be corrected by a mechanism called mismatch repair. While the error rate of DNA replication is about one in \(10^7\) nucleotides in the absence of mismatch repair, this rate is further reduced a hundred-fold to one in \)10^9\) nucleotides when mismatch repair is functional.
What are the tasks that a mismatch repair system must accomplish?
- Scan newly made DNA to see if there are any mispaired bases (e.g., a G paired to a T)
- Identify and cut out the region of the mismatch
- Correctly fill in the gap created by the excision of the mismatch region.
Importantly, the mismatch repair system must have a means to distinguish the newly made DNA strand from the template strand, if replication errors are to be fixed correctly. In other words, when the mismatch repair system encounters an A-G mispair, for example, it must know whether the A should be removed and replaced with a C or if the G should be removed and replaced with a T.
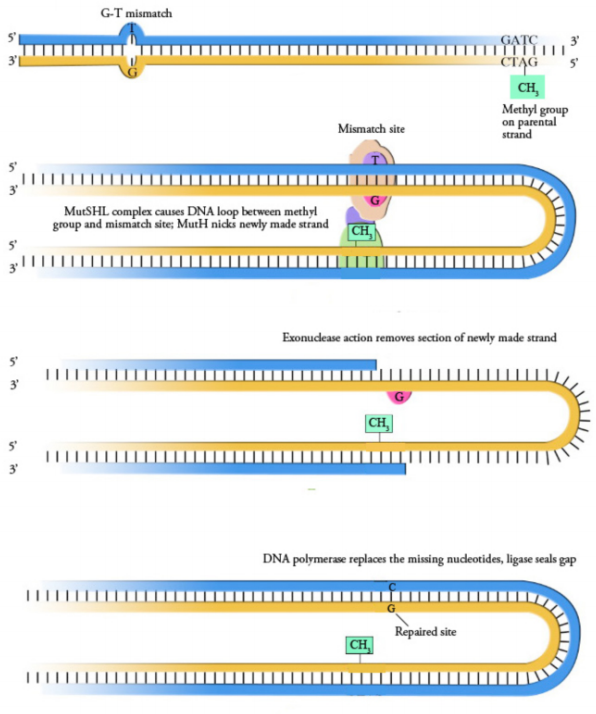
Mismatch repair has been well studied in bacteria, and the proteins involved have been identified. Eukaryotes have a mismatch repair system that repairs not only single base mismatches but also insertions and deletions. In bacteria, mismatch repair proteins are encoded by a group of genes collectively known as the Mut (for mutator and pronounced "mute") genes. Some of the most important components of the mismatch repair machinery are the proteins MutS, MutL, and MutH (sometimes known as MutSHL complex). MutS recognizes the mismatch and recruits MutL and MutH, as well as other proteins which help cut out the region containing the mismatch. A DNA polymerase and ligase fill in the gap and join the ends, respectively.
But how does the mismatch repair system distinguish between the original and the new strands of DNA? Bacteria have a system that methylates DNA at GATC sequences to solve this problem. E.coli has an enzyme that adds methyl groups on the to adenines in GATC sequences. Newly-replicated DNA strands lacks this methylation and thus, can be distinguished from the template strand, which is methylated. The Mut genes use the methylation to direct removal of the non-methylated strand, keeping the methylated strand as the template to repair the region. Because methylation is the criterion that enables the mismatch repair system to choose the strand that is repaired, the bacterial mismatch repair system is described as being methyl-directed. Note that the newly-synthesized strand will eventually also be methylated by the cellular enzymes, so this repair system is only useful immediately following DNA replication, before the new strand becomes marked as well.
Although eukaryotic cells have Mut gene homologs, they do not use the same methylation mechanism to distinguish the new strand from the template. Although the mechanisms, are not yet fully understood, recent research suggests that the mismatch repair system in eukaryotes may be coupled to DNA replication so that the strand being synthesized strand can be identified via nicks or proteins asymmetrically bound to the replication machinery (reviewed in Reyes et al. 2015).
Systems to Repair Damage to DNA
Some mistakes are made when DNA is copied, if the wrong base is inserted during synthesis of the new strand. But even DNA that is not being replicated can get damaged or mutated. These sorts of damage are not associated with DNA replication, rather they can occur at any time.
Recall that some major causes of DNA damage are:
- Radiation (e.g., UV rays in sunlight, in tanning booths)
- Exposure to damaging chemicals (such as benzopyrene in car exhaust and cigarette smoke)
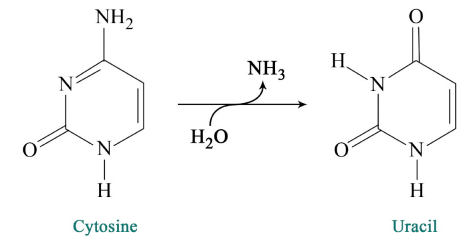
- Chemical reactions within the cell (such as the deamination of cytosine to give uracil).
How do cells repair such damage? Cells have several ways to remove the sorts of damage described above, with excision repair being a common strategy. Excision repair is a general term for the cutting out and re-synthesis of the damaged region of the DNA. There are a couple of varieties of excision repair:
Direct repair
Cells may possess mechanisms to repair types of damage that occur frequently, for example there are proteins that remove alkyl groups from DNA bases. Most organisms, especially those exposed to high amounts of UV radiation, have enzymes that can directly repair thymine dimers. The process, called photoreactivation or light repair, uses an enzyme, photolyase, that depends on visible light to directly breaks the bonds joining the thymine bases. Notably, placental mammals do not have photolyases capable of this type of repair and so must use an alternate pathway to repair UV-induced thymine dimers.
Nucleotide Excision Repair (NER)
This system fixes damage by chemicals as well as UV damage. In nucleotide excision repair, the damage is recognized and a cut is made on either side of the damaged region by an endonuclease enzyme (sometimes called an excision nuclease or exinuclease). A short portion of the DNA strand containing the damage is then removed and a DNA polymerase fills in the gap with the appropriate nucleotides. The newly made DNA is joined to the rest of the DNA backbone by the enzyme DNA ligase. In E. coli, NER is carried out by a group of proteins encoded by the uvrABC genes. As you can see, NER is similar, in principle, to mismatch repair. However, in NER, the distortion of the helix, caused by the DNA damage, clearly indicates which strand of the DNA needs to be removed and replaced.
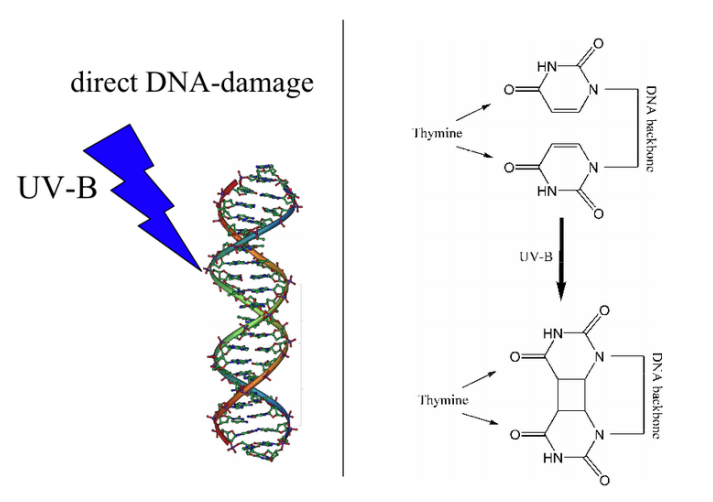
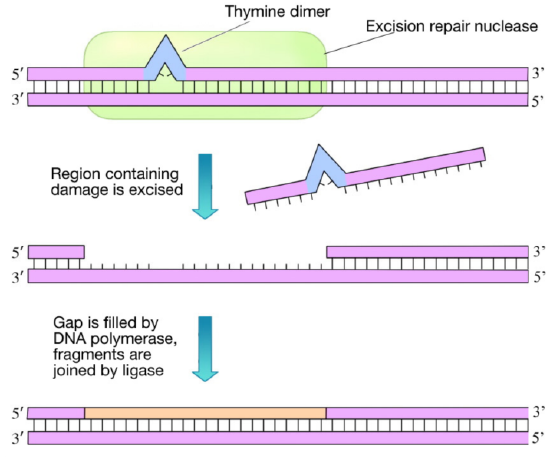
Figure \(\PageIndex{3}\): (Left) Thymine dimer formation can occur when UV-B light hits DNA and covalent bonds form between adjacent thymine bases. (Right) Thymine dimers can be removed by an excision repair nuclease that recognizes the dimer, excising the region of one strand of DNA containing the dimer. The gap is filled by DNA polymerase and joined to the original strand by ligase.
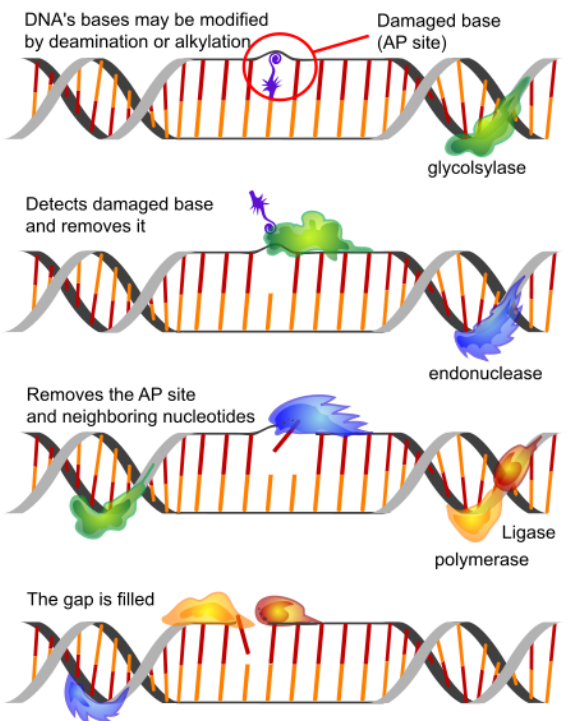
Base Excision Repair (BER)
BER deals with situations like the deamination of cytosine to uracil. As noted earlier, cytosines in DNA sometimes undergo deamination to form the base uracil. Because cytosines pair with guanines and uracils pair with adenine, the conversion of cytosine to uracil in the DNA would lead to the insertion of an A in the newly replicated strand instead of the G that should have gone in across from a C. To prevent this from happening, uracils are removed from DNA by base excision repair.
In base excision repair, a single base is first removed from the DNA, followed by removal of a region of the DNA surrounding the missing base. The gap is then repaired. The removal of uracil from DNA is accomplished by the enzyme uracil DNA glycosylase, which breaks the bond between the uracil and the sugar in the nucleotide. The removal of the uracil base creates a gap called an apyrimidinic site (AP site). The presence of the AP site triggers the activity of an AP endonuclease that cuts the DNA backbone. A short region of the DNA surrounding the site of the original uracil is then removed and replaced.
Nobel Prize Winning Work
Tomas Lindahl, Paul Modrich and Aziz Sancar shared the 2015 Nobel Prize in Chemistry "for mechanistic studies of DNA repair" for their characterization of the proteins and processes involved in mismatch repair, base excision repair, and nucleotide excision repair (https://www.nobelprize.org/prizes/chemistry/2015/press-release/).
Additional repair pathways
What if DNA damage is not isolated to the bases? Damage to DNA may also involved breakage of the sugar-phosphate backbone of the chromosome. Cells have some additional options to repair damage to large regions of DNA. The first is non-homologous end-joining (NHEJ). As the name suggests, in this process the ends of two broken chromosomes are ligated together without idenityfing whether or not they were actually part of the same chromosome. This mechanism can be thought of as an emergency response to broken chromosomes; without repair the broken ends of DNA could be further damaged by exonucleases, so joining the two ends prevents further loss of genetic material. However, the process is imperfect and can result in insertions, deletions, or translocations (joining of two different chromosomes). A second option is homologous recombination repair, in which a homologous chromosome is used as a template to synthesize a broken or damaged region of DNA. The fundamental mechanism here is similar to the process of recombination used during meiosis.
Contributors and Attributions
Dr. Kevin Ahern and Dr. Indira Rajagopal (Oregon State University)
References
Reyes, G.X., Schmidt, T.T., Kolodner, R.D. et al. New insights into the mechanism of DNA mismatch repair. Chromosoma 124, 443–462 (2015). https://doi.org/10.1007/s00412-015-0514-0