
2.3.4: C4. Common Motifs in Proteins
- Page ID
- 64216

Super-Secondary Structure - Given the number of possible combinations of 1°, 2°, and 3° structures, one might guess that the 3D structure of each protein is quite distinctive. This is true. However, it has been found that similar substructures are found in proteins. For instance, common secondary structures are often grouped together to form a motifs called super-secondary structure (SSS). See some examples below:
- helix-loop-helix : found in DNA binding proteins and also in calcium binding proteins. This motif, which is also a helix-loop-helix, is often called the EF hand. The loop region in calcium binding proteins are enriched in Asp, Glu, Ser, and Thr. Why? The EF hand shown below is from calmodulin.
Figure: helix-loop-helix (image made with VMD)

Figure: EF Hand

 Jmol: Updated helix-loop-helix of the lambda Repressor Jmol14 (Java) | JSMol (HTML5)
Jmol: Updated helix-loop-helix of the lambda Repressor Jmol14 (Java) | JSMol (HTML5)
Jmol: Updated helix-loop-helix (EF hand) from calmodulin Jmol14 (Java) | JSMol (HTML5)
- beta-hairpin or beta-beta: is present in most antiparallel beta structures both as an isolated ribbon and as part of beta sheets.
Figure: beta-hairpin, or beta-beta (image made with VMD)

Jmol: Updated beta-hairpin from bovine pancreatic trypsin inhibitor Jmol14 (Java) | JSMol (HTML5)
- Greek Key motif: four adjacent antiparallel beta strands are often arranged in a pattern similar to the repeating unit of one of the ornamental patterns used in ancient Greece.
Figure: Greek Key Motiff

Jmol: Greek Key
- Figure: beta-alpha-beta: is a common way to connect two parallel beta strands. (beta hairpin used for antiparallel beta strands).
Figure: beta-alpha-beta (image made with VMD with H atoms added by Molprobity

Jmol: Updated beta-helix-beta motif from triose phosphate isomerase Jmol14 (Java) | JSMol (HTML5)
- Beta Helices: These right-handed parallel helix structures consists of a contiguous polypeptide chain with three parallel beta strands separated by three turns forming a single rung of a larger helical structure which in total might contain as many as nine rungs. The intrastrand H-bonds are between parallel beta strands in separate rungs. These seem to prevalent in pathogens (bacteria, viruses, toxins) proteins that facilitate binding of the pathogen to a host cell.
Figure: Beta Helices (image made with VMD)
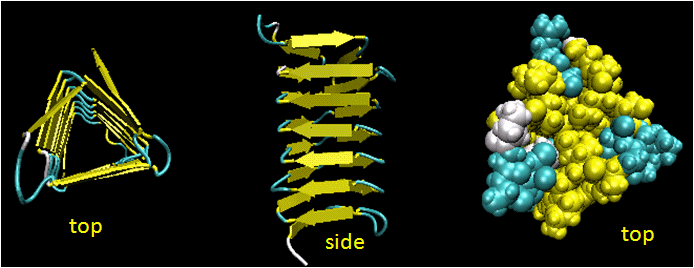
Table: Beta Helices Vibrio cholerae cholera Helicobacter pylori ulcers Plasmodium falciparum malaria Chlamyidia trachomatis VD Chlamydophilia pneumoniae respiratory infection Trypanosoma brucei sleeping sickness Borrelia burgdorferi Lymes disease Bordetella parapertussis whooping cough Bacillus anthracis anthrax Neisseria meningitides menigitis Legionaella pneumophilia Legionaire's disease  Beta Topologies on the Web
Beta Topologies on the Web
- of the Swiss Institute of Bioinformatics. (SIB) is dedicated to the analysis of protein sequences and structures as well as 2-D PAGE
Domains -
Domains are the fundamental unit of 3o structure. It can be considered a chain or part of a chain that can independently fold into a stable tertiary structure. Domains are units of structure but can also be units of function. Some proteins can be cleaved at a single peptide bonds to form two fragments. Often, these can fold independently of each other, and sometimes each unit retains an activity that was present in the uncleaved protein. Sometimes binding sites on the proteins are found in the interface between the structural domains. Many proteins seem to share functional and structure domains, suggesting that the DNA of each shared domain might have arisen from duplication of a primordial gene with a particular structure and function.
Evolution has led towards increasing complexity which has required proteins of new structure and function. Increased and different functionalities in proteins have been obtained with additions of domains to base protein. Chothia (2003) has defined domain in an evolutionary and genetic sense as "an evolutionary unit whose coding sequence can be duplicated and/or undergo recombination". Proteins range from small with a single domain (typically from 100-250 amino acids) to large with many domains. From recent analyzes of genomes, new protein functionalities appear to arise from addition or exchange of other domains which, according to Chothia, result from
- "duplication of sequences that code for one or more domains
- divergence of duplicated sequences by mutations, deletions, and insertions that produce modified structures that may have useful new properties to be selected
- recombination of genes that result in novel arrangement of domains."
Structural analyses show that about half of all protein coding sequences in genomes are homologous to other known protein structures. There appears to be about 750 different families of domains (i.e small proteins derived from a common ancestor) in vertebrates, each with about 50 homologous structures. About 430 of these domain families are found in all the genomes that have been solved.