
5.1: Introduction to Enumeration of Bacteria
- Page ID
- 52279
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Learning Outcomes
- Introduction to dilution theory
- Estimate the number of microbes in a sample using serial dilution techniques and standard/viable plate counts
Enumeration of Bacteria
Often one needs to determine the number of organisms in a sample of material, for example, in water, foods, or a bacterial culture. For example, bacterial pathogens can be introduced into foods at any stage: during growth/production at the farm, during processing, during handling and packaging, and when the food is prepared in the kitchen (1). In general, small numbers of pathogenic bacteria are not dangerous, but improper storage and/or cooking conditions can allow these bacteria to multiply to dangerous levels (1).
Fecal contamination of water is another one of the ways in which pathogens can be introduced (1). Coliform bacteria are Gram-negative non-spore forming bacteria that are capable of fermenting lactose to produce acid and gas. A subset of these bacteria are the fecal coliforms, which are found at high levels in human and animal intestines. Fecal coliform bacteria such as E. coli, are often used as indicator species, as they are not commonly found growing in nature in the absence of fecal contamination (1). The presence of E. coli suggests feces are present, indicating that serious pathogens, such as Salmonella species and Campylobacter species, could also be present (1).
Methods of Enumeration
Many approaches are commonly employed for enumerating bacteria, including measurements of the direct microscopic count, culture turbidity, dry weight of cells, etc. In a microbiology lab, we frequently determine the total viable count in a bacterial culture.
The most common method of measuring viable bacterial cell numbers is the standard or viable plate count or colony count. This is a viable count, NOT a total cell count. It reveals information related only to viable or live bacteria. Using this method, a small volume (0.1 - 1.0 mL) of liquid containing an unknown number of bacteria is spread over the surface of an agar plate, creating a "spread plate." The spread plates are incubated for 24-36 hours. During that time, each individual viable bacterial cell multiplies to form a readily visible colony. The number of colonies is then counted and this number should equal the number of viable bacterial cells in the original volume of sample, which was applied to the plate.
For accurate information, it is critical that each colony comes from only one cell, so chains and clumps of cells must be broken apart. However, many bacterial species grow in pairs, chains, or clusters, or they may have sticky capsules or slime layers, which cause them to clump together. It is sometimes difficult to separate these into single cells, which in turn makes it difficult to obtain an accurate count of the original cell numbers. Therefore, the total number of viable cells obtained from this procedure is usually reported as the number of colony-forming units (CFUs).
A bacterial culture and many other samples usually contain too many cells to be counted directly. Thus, in order to obtain plates, which are not hopelessly overgrown with colonies, it is often necessary to dilute the sample and spread measured amounts of the diluted sample on plates. Dilutions are performed by careful aseptic pipetting of a known volume of sample into a known volume of a sterile buffer or sterile water. This is mixed well and can be used for plating and/or further dilution. If the number of cells in the original sample is unknown, then a wide range of dilutions are usually prepared and plated. The preparation of dilutions and the calculation and use of dilution factors to obtain the number of microorganisms present in a sample are important basic techniques in microbiology.
Method:
Aliquots from a stepwise or serial dilution of the original sample are spread on plates. Only a few of the plates following incubation will contain a suitable number of colonies to count; those plated from low dilutions may contain too many colonies to count easily while those plated from high dilutions may contain too few colonies or none at all. Ideally plates containing 30-300 colonies per plate should be counted. At this colony number, the number counted is high enough to have statistical accuracy, yet low enough to avoid mistakes due to overlapping colonies.

Figure 1: Serial dilution series and plating. A wide series of dilutions (e.g. 10-2 to 10-8 ) is normally performed on the sample culture and spread plates created from the dilutions. A number of spread plates is needed because the exact number of live bacteria in the sample is usually unknown. Greater accuracy can be achieved by plating duplicates or triplicates of each dilution.

Image 1: Picture of spread plates showing bacterial growth (Escherichia coli, 40 hours, 25°C) on five plates prepared from a ten-fold dilution series. Care was taken to avoid spreading to the edges of the plates as it is more difficult to count colonies along the edge of the agar. Note how many colonies are in the plate from the 10-1 & 10-2 dilution plates. These plates have densely packed colonies, are too numerous to count, and most likely more than 300 CFUs. On the other hand, plates 10-3 , 10-4 and 10-5 have a countable number (between 30-300) of CFUs. Image by Kathryn Wise and Darel Paulson, Minnesota State University, Moorhead, MN.
Calculations
In order to make the calculation of the number of cells/mL in the original sample less formidable, dilutions are designed to be easy to handle mathematically. The most common dilutions are ten-fold and multiples of ten-fold. A 1/10 or 10-1 dilution can be achieved by mixing 1 mL of sample with 9 mL of sterile dilution buffer. A subsequent 1/10 dilution of this first ten-fold dilution, made by mixing 1 mL of this first dilution with 9 mL of fresh sterile dilution buffer, would give a total dilution of the original sample of 100-fold (1/10 X 1/10 = 1/100 or 10-1 X 10-1 = 10-2). Alternatively, a 100-fold dilution can be made directly from the original sample by mixing 1 mL of sample and 99 mL of buffer. These dilutions can be made in successive steps or a series to give a wide range for any given sample.
Once the dilution is made, an aliquot can be spread on an agar plate to create a spread plate. After incubation, the colonies which arise can be counted and the number of cells (more precisely the number of colony-forming units or CFUs) in the original sample can be calculated. For example in the image below, you did a serial dilution of a culture of the red pigmented bacterium, Serratia marcescens and made a series of spread plates. In plate 1, this was the 10-1 dilution, in plate 2 is the 10-2 dilution, and in plate 3 is the 10-3 dilution. After incubation, you count 241 colonies were present on the plate 10-2 dilution.
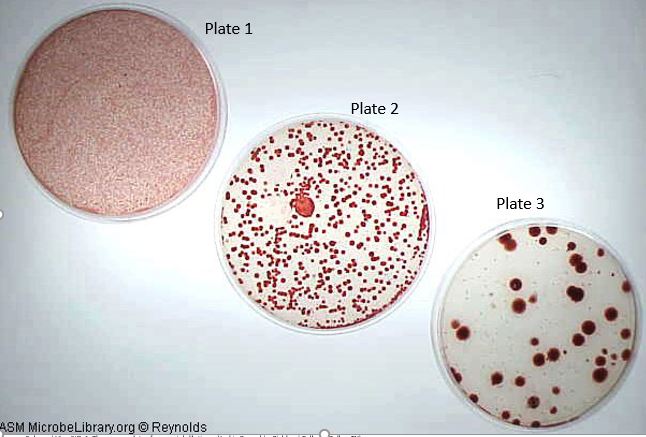
Image 2: Three spread plates from serial dilutions. Image by Jackie Reynolds, Richland College, Dallas, TX.
Therefore, there were 241 X 102 CFU/mL in the original sample or expressed 2.41 x 104. To arrive at this final number you only need to multiply the final number of colonies on the plate, 241, times the total dilution factor. The dilution factor is defined as the inverse of the dilution. So in this case, the dilution factor is the inverse of 1/100 or 100. In other words, the dilution factor is how many times the sample was diluted.
It usually works better to spread only 0.1 mL of a sample on a standard-size petri plate. If the above example were changed such that 0.1 mL of a 100-fold dilution of the same sample was plated, there would ideally have been 24 colonies on the plate. This number represents the number of CFU in only 0.1 mL of the dilution plated. Therefore, to calculate the CFU/ml in the sample it is necessary to multiply the number of colonies on the plate by 10 (there are ten 0.1 mL units in 1.0 mL) and then by the dilution factor (100) to arrive at the final answer: 24 CFU x 10 x 100 = 24000 or 2.4 x 104 CFU/mL.
The only way to understand dilution theory well is to practice it, so you should work practice problems until you feel confident in using dilution factors and calculating CFU/mL in original samples. You should also be able to determine the proper dilutions to use to obtain 30-300 colonies on a plate if the original number of CFU/mL in a sample is known.
Watch Video 1 on how to perform a serial dilution using "pour plates" and the associated calculations:
Watch Video 1: Serial dilutions and pour plate technique. The example here using a "pour plate" technique to spread the dilutions out instead of the "spread plate" discussed above, but the outcome of both techniques of spreading the dilution sample out is the same. (10:48) Video by Microbial Zoo. URL:https://www.youtube.com/watch?v=nViO9Y4Yxfk
Watch video 2 on how to perform a serial dilution and make spread plates.
Watch Video 2: Serial dilutions with a bacterial sample and the 'spread plating' technique at NC State Microbiology labs. (10:30) URL: https://youtu.be/IJcw4fRsYnU
References
1. Contributed by Joan Petersen & Susan McLaughlin Associate Professors (Biological Sciences and Geology) at Queensborough Community College