
3.3: Evidence of Evolution
- Page ID
- 32473

The evidence for evolution is compelling and extensive. Looking at every level of organization in living systems, biologists see the signature of past and present evolution. Darwin dedicated a large portion of his book, On the Origin of Species, identifying patterns in nature that were consistent with evolution and since Darwin our understanding has become clearer and broader.
Fossils
Fossils provide solid evidence that organisms from the past are not the same as those found today; fossils show a progression of evolution. Scientists determine the age of fossils and categorize them all over the world to determine when the organisms lived relative to each other. The resulting fossil record tells the story of the past, and shows the evolution of form over millions of years (Figure \(\PageIndex{1}\)). For example, highly detailed fossil records have been recovered for sequences of species in the evolution of whales and modern horses. The fossil record of horses in North America is especially rich and many contain transition fossils: those showing intermediate anatomy between earlier and later forms. The fossil record extends back to a dog-like ancestor some 55 million years ago that gave rise to the first horse-like species 55 to 42 million years ago in the genus Eohippus. The series of fossils tracks the change in anatomy resulting from a gradual drying trend that changed the landscape from a forested one to a prairie. Successive fossils show the evolution of teeth shapes and foot and leg anatomy to a grazing habit, with adaptations for escaping predators, for example in species of Mesohippus found from 40 to 30 million years ago. Later species showed gains in size, such as those of Hipparion, which existed from about 23 to 2 million years ago. The fossil record shows several adaptive radiations in the horse lineage, which is now much reduced to only one genus, Equus, with several species.

Anatomy and Embryology
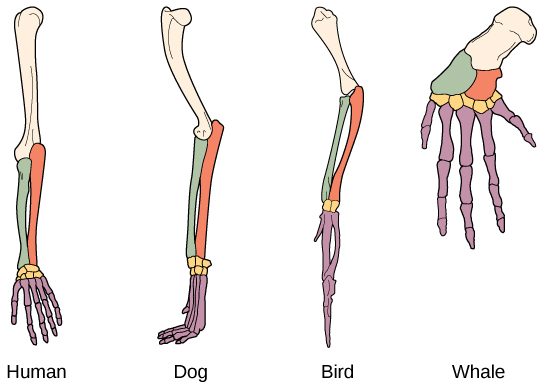
Another type of evidence for evolution is the presence of structures in organisms that share the same basic form. For example, the bones in the appendages of a human, dog, bird, and whale all share the same overall construction (Figure \(\PageIndex{2}\)). That similarity results from their origin in the appendages of a common ancestor. Over time, evolution led to changes in the shapes and sizes of these bones in different species, but they have maintained the same overall layout, evidence of descent from a common ancestor. Scientists call these synonymous parts homologous structures. Some structures exist in organisms that have no apparent function at all, and appear to be residual parts from a past ancestor. For example, some snakes have pelvic bones despite having no legs because they descended from reptiles that did have legs. These unused structures without function are called vestigial structures. Other examples of vestigial structures are wings on flightless birds (which may have other functions), leaves on some cacti, traces of pelvic bones in whales, and the sightless eyes of cave animals.
Another evidence of evolution is the convergence of form in organisms that share similar environments. For example, species of unrelated animals, such as the arctic fox and ptarmigan (a bird), living in the arctic region have temporary white coverings during winter to blend with the snow and ice (Figure \(\PageIndex{3}\)). The similarity occurs not because of common ancestry, indeed one covering is of fur and the other of feathers, but because of similar selection pressures—the benefits of not being seen by predators.

Embryology, the study of the development of the anatomy of an organism to its adult form also provides evidence of relatedness between now widely divergent groups of organisms. Structures that are absent in some groups often appear in their embryonic forms and disappear by the time the adult or juvenile form is reached. For example, all vertebrate embryos, including humans, exhibit gill slits at some point in their early development. These disappear in the adults of terrestrial groups, but are maintained in adult forms of aquatic groups such as fish and some amphibians. Great ape embryos, including humans, have a tail structure during their development that is lost by the time of birth. The reason embryos of unrelated species are often similar is that mutational changes that affect the organism during embryonic development can cause amplified differences in the adult, even while the embryonic similarities are preserved.
Biogeography
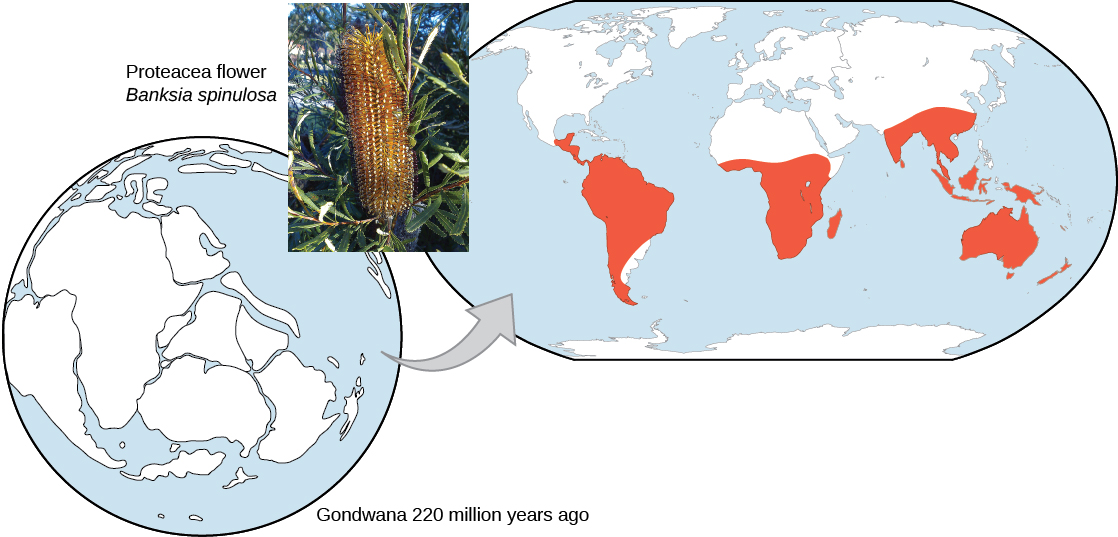
The geographic distribution of organisms on the planet follows patterns that are best explained by evolution in conjunction with the movement of tectonic plates over geological time. Broad groups that evolved before the breakup of the supercontinent Pangaea (about 200 million years ago) are distributed worldwide. Groups that evolved since the breakup appear uniquely in regions of the planet, for example the unique flora and fauna of northern continents that formed from the supercontinent Laurasia and of the southern continents that formed from the supercontinent Gondwana. The presence of Proteaceae in Australia, southern Africa, and South America is best explained by the plant family’s presence there prior to the southern supercontinent Gondwana breaking up (Figure \(\PageIndex{4}\)).
The great diversification of the marsupials in Australia and the absence of other mammals reflects that island continent’s long isolation. Australia has an abundance of endemic species—species found nowhere else—which is typical of islands whose isolation by expanses of water prevents migration of species to other regions. Over time, these species diverge evolutionarily into new species that look very different from their ancestors that may exist on the mainland. The marsupials of Australia, the finches on the Galápagos, and many species on the Hawaiian Islands are all found nowhere else but on their island, yet display distant relationships to ancestral species on mainlands.
Molecular Biology
Like anatomical structures, the structures of the molecules of life reflect descent with modification. Evidence of a common ancestor for all of life is reflected in the universality of DNA as the genetic material and of the near universality of the genetic code and the machinery of DNA replication and expression. Fundamental divisions in life between the three domains are reflected in major structural differences in otherwise conservative structures such as the components of ribosomes and the structures of membranes. In general, the relatedness of groups of organisms is reflected in the similarity of their DNA sequences—exactly the pattern that would be expected from descent and diversification from a common ancestor.
DNA sequences have also shed light on some of the mechanisms of evolution. For example, it is clear that the evolution of new functions for proteins commonly occurs after gene duplication events. These duplications are a kind of mutation in which an entire gene is added as an extra copy (or many copies) in the genome. These duplications allow the free modification of one copy by mutation, selection, and drift, while the second copy continues to produce a functional protein. This allows the original function for the protein to be kept, while evolutionary forces tweak the copy until it functions in a new way.
Section Summary
The evidence for evolution is found at all levels of organization in living things and in the extinct species we know about through fossils. Fossils provide evidence for the evolutionary change through now extinct forms that led to modern species. For example, there is a rich fossil record that shows the evolutionary transitions from horse ancestors to modern horses that document intermediate forms and a gradual adaptation to changing ecosystems. The anatomy of species and the embryological development of that anatomy reveal common structures in divergent lineages that have been modified over time by evolution. The geographical distribution of living species reflects the origins of species in particular geographic locations and the history of continental movements. The structures of molecules, like anatomical structures, reflect the relationships of living species and match patterns of similarity expected from descent with modification.
Glossary
- vestigial structure
- a physical structure present in an organism but that has no apparent function and appears to be from a functional structure in a distant ancestor
Contributors and Attributions
Samantha Fowler (Clayton State University), Rebecca Roush (Sandhills Community College), James Wise (Hampton University). Original content by OpenStax (CC BY 4.0; Access for free at https://cnx.org/contents/b3c1e1d2-83...4-e119a8aafbdd).