
8: Introduction to Viruses
- Page ID
- 10660

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Viruses are typically described as obligate intracellular parasites, acellular infectious agents that require the presence of a host cell in order to multiply. Viruses that have been found to infect all types of cells – humans, animals, plants, bacteria, yeast, archaea, protozoa…some scientists even claim they have found a virus that infects other viruses! But that is not going to happen without some cellular help.
Virus Characteristics
Viruses can be extremely simple in design, consisting of nucleic acid surrounded by a protein coat known as a capsid. The capsid is composed of smaller protein components referred to as capsomers. The capsid+genome combination is called a nucleocapsid.
Viruses can also possess additional components, with the most common being an additional membranous layer that surrounds the nucleocapsid, called an envelope. The envelope is actually acquired from the nuclear or plasma membrane of the infected host cell, and then modified with viral proteins called peplomers. Some viruses contain viral enzymes that are necessary for infection of a host cell and coded for within the viral genome. A complete virus, with all the components needed for host cell infection, is referred to as a virion.

Virus Characteristics, Image created by Ben Taylor, Public Domain, Via Wikipedia commons
Virus Genome
While cells contain double-stranded DNA for their genome, viruses are not limited to this form. While there are dsDNA viruses, there are also viruses with single-stranded DNA (ssDNA), double-stranded RNA (dsRNA), and single-stranded RNA (ssRNA). In this last category, the ssRNA can either positive-sense (+ssRNA, meaning it can transcribe a message, like mRNA) or it can be negative-sense (-ssRNA, indicating that it is complementary to mRNA). Some viruses even start with one form of nucleic acid in the nucleocapsid and then convert it to a different form during replication.
Virus Structure
Viral nucleocapsids come in two basic shapes, although the overall appearance of a virus can be altered by the presence of an envelope, if present. Helical viruses have an elongated tube-like structure, with the capsomers arranged helically around the coiled genome. Icosahedral viruses have a spherical shape, with icosahedral symmetry consisting of 20 triangular faces. The simplest icosahedral capsid has 3 capsomers per triangular face, resulting in 60 capsomers for the entire virus. Some viruses do not neatly fit into either of the two previous categories because they are so unusual in design or components, so there is a third category known as complex viruses. Examples include the poxvirus with a brick-shaped exterior and a complicated internal structure, as well as bacteriophage with tail fibers attached to an icosahedral head.
Virus Replication Cycle
While the replication cycle of viruses can vary from virus to virus, there is a general pattern that can be described, consisting of five steps:
- Attachment – the virion attaches to the correct host cell.
- Penetration or Viral Entry – the virus or viral nucleic acid gains entrance into the cell.
- Synthesis – the viral proteins and nucleic acid copies are manufactured by the cells’ machinery.
- Assembly – viruses are produced from the viral components.
- Release – newly formed virions are released from the cell.
Attachment
Outside of their host cell, viruses are inert or metabolically inactive. Therefore, the encounter of a virion to an appropriate host cell is a random event. The attachment itself is highly specific, between molecules on the outside of the virus and receptors on the host cell surface. This accounts for the specificity of viruses to only infect particular cell types or particular hosts.
Penetration or Viral Entry
Many unenveloped (or naked) viruses inject their nucleic acid into the host cell, leaving an empty capsid on the outside. This process is termed penetration and is common with bacteriophage, the viruses that infect bacteria. With the eukaryotic viruses, it is more likely for the entire capsid to gain entrance into the cell, with the capsid being removed in the cytoplasm. An unenveloped eukaryotic virus often gains entry via endocytosis, where the host cell is compelled to engulf the capsid resulting in an endocytic vesicle. An enveloped eukaryotic virus gains entrance for its nucleocapsid when the viral envelope fuses with the host cell membrane, pushing the nucleocapsid past the cell membrane. If the entire nucleocapsid is brought into the cell then there is an uncoating process to strip away the capsid and release the viral genome.
Synthesis
The synthesis stage is largely dictated by the type of viral genome, since genomes that differ from the cell’s dsDNA genome can involve intricate viral strategies for genome replication and protein synthesis. Viral specific enzymes, such as RNA-dependent RNA polymerases, might be necessary for the replication process to proceed. Protein production is tightly controlled, to insure that components are made at the right time in viral development.
Assembly
The complexity of viral assembly depends upon the virus being made. The simplest virus has a capsid composed of 3 different types of proteins, which self-assembles with little difficulty. The most complex virus is composed of over 60 different proteins, which must all come together in a specific order. These viruses often employ multiple assembly lines to create the different viral structures and then utilize scaffolding proteins to put all the viral components together in an organized fashion.
Release
The majority of viruses lyse their host cell at the end of replication, allowing all the newly formed virions to be released to the environment. Another possibility, common for enveloped viruses, is budding, where one virus is released from the cell at a time. The cell membrane is modified by the insertion of viral proteins, with the nucleocapsid pushing out through this modified portion of the membrane, allowing it to acquire an envelope.
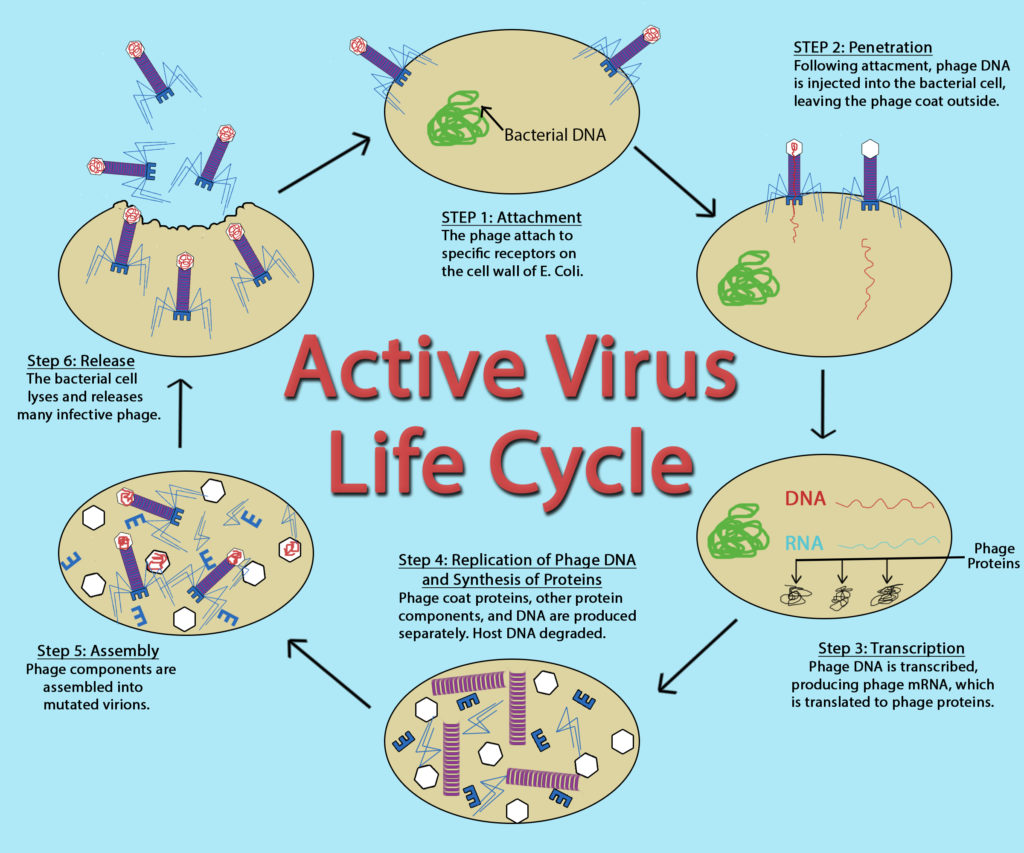
Active Virus Life Cycle by John Kellogg Via OER at Oregon State University
Bacteriophage
Viruses that infect bacteria are known as bacteriophage or phage. A virulent phage is one that always lyses the host cell at the end of replication, after following the five steps of replication described above. This is called the lytic cycle of replication.
There are also temperate phage, viruses that have two options regarding their replication. Option 1 is to mimic a virulent phage, following the five steps of replication and lysing the host cell at the end, referred to as the lytic cycle. But temperate phage differ from virulent phage in that they have another choice: Option 2, where they remain within the host cell without destroying it. This process is known as lysogeny or the lysogenic cycle of replication.
A phage employing lysogeny still undergoes the first two steps of a typical replication cycle, attachment and penetration. Once the viral DNA has been inserted into the cell it integrates with the host DNA, forming a prophage. The infected bacterium is referred to as a lysogenor lysogenic bacterium. In this state, the virus enjoys a stable relationship with its host, where it does not interfere with host cell metabolism or reproduction. The host cell enjoys immunity from reinfection from the same virus.
Exposure of the host cell to stressful conditions (i.e. UV irradiation) causes induction, where the viral DNA excises from the host cell DNA. This event triggers the remaining steps of the lytic cycle, synthesis, maturation, and release, leading to lysis of the host cell and release of newly formed virions.
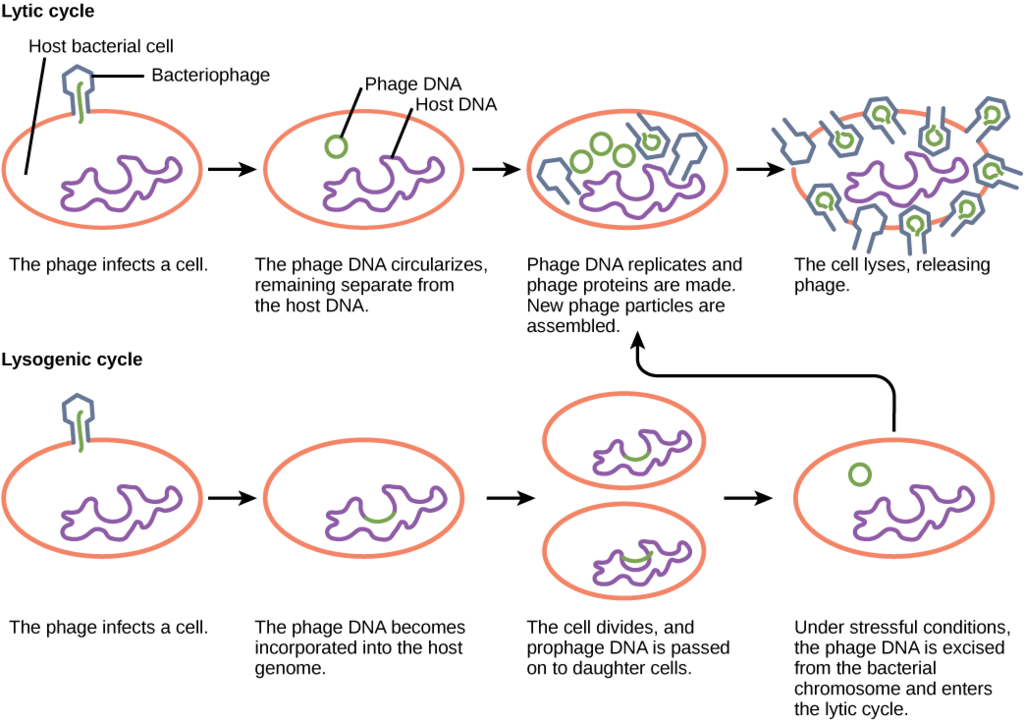
Lytic Cycle Versus Lysogenic Cycle of Replication. OpenStax, Virus Infections and Hosts. OpenStax CNX. Apr 11, 2013 http://cnx.org/contents/7cbd15ad-5bff-4678-a99f-85fd579e070c@3.
So, what dictates the replication type that will be used by a temperate phage? If there are plenty of host cells around, it is likely that a temperate phage will engage in the lytic cycle of replication, leading to a large increase in viral production. If host cells are scarce, a temperate phage is more likely to enter lysogeny, allowing for viral survival until host cell numbers increase. The same is true if the number of phage in an environment greatly outnumber the host cells, since lysogeny would allow for host cells numbers to rebound, ensuring long term viral survival.
Lysogens can experience a benefit from lysogeny as well, since it can result in lysogenic conversion, a situation where the development of a prophage leads to a change in the host’s phenotype. One of the best examples of this is for the bacterium Corynebacterium diphtheriae, the causative agent of diphtheria. The diphtheria toxin that causes the disease is encoded within the phage genome, so only C. diphtheriaelysogens cause diphtheria.
Eukaryotic Viruses
Eukaryotic viruses can cause one of four different outcomes for their host cell. The most common outcome is host cell lysis, resulting from a virulent infection (essentially the lytic cycle of replication seen in phage). Some viruses can cause a latent infection, co-existing peacefully with their host cells for years (much like a temperate phage during lysogeny). Some enveloped eukaryotic viruses can also be released one at a time from an infected host cell, in a type of budding process, causing a persistent infection. Lastly, certain eukaryotic viruses can cause the host cell to transform into a malignant or cancerous cell, a mechanism known as transformation.
Viruses and Cancer
There are many different causes of cancer, or unregulated cell growth and reproduction. Some known causes include exposure to certain chemicals or UV light. There are also certain viruses that have a known associated with the development of cancer. Such viruses are referred to as oncoviruses. Oncoviruses can cause cancer by producing proteins that bind to host proteins known as tumor suppressor proteins, which function to regulate cell growth and to initiate programmed cell death, if needed. If the tumor suppressor proteins are inactivated by viral proteins then cells grow out of control, leading to the development of tumors and metastasis, where the cells spread throughout the body.
Key Words
virus, obligate intracellular parasite, capsid, bacteriophage, capsomere, nucleocapsid, envelope, peplomer, virion, dsDNA, ssDNA, dsRNA, +ssRNA, -ssRNA, helical viruses, icosahedral viruses, complex viruses, attachment, penetration, viral entry, synthesis, assembly, release, naked virus, endocytosis, budding, bacteriophage, phage, virulent phage, lytic cycle, temperate phage, lysogeny, lysogenic cycle, prophage, lysogen, lysogenic bacterium, induction, lysogenic conversion, virulent infection, latent infection, persistent infection, transformation, oncovirus, tumor suppressor proteins.
Essential Questions/Objectives
- What are the general properties of a virus?
- What is the size range of viruses? How do they compare, size-wise, to bacteria?
- What is the general structure of viruses? What are the different components?
- What viral shapes exist?
- How do envelopes and enzymes relate to viruses?
- What types of viral genomes exist?
- What are the steps of viral multiplication? What is happening at each step? How do Bacterial/Archaeal viruses differ from eukaryotic viruses, in regards to multiplication?
- What are the 2 types of viral infection found in Bacteria/Archaea? What are the specific terms associated with viral infection of bacterial/archaeal cells?
- What are the 4 types of viral infection found in eukaryotes?
- How do some viruses cause cancer?
Exploratory Questions (OPTIONAL)
- What is the largest bacterium or archaean ever discovered? What is the smallest eukaryote ever discovered?