
15.6: Mitosis
- Page ID
- 16189

Mitosis consists of prophase, metaphase, anaphase, and telophase, with distinct cellular activities characterizing each phase. This completes the duplication of the nucleus, and is followed by cytokinesis, in which the cell divides to produce two daughter cells.
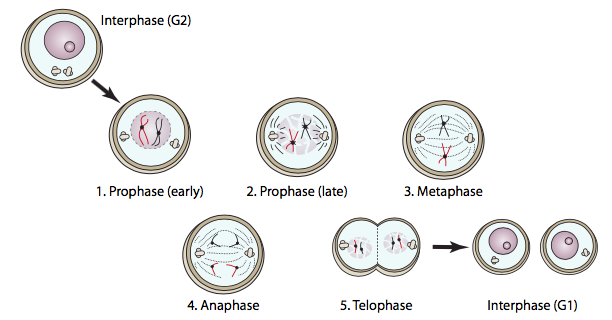
Prophase is the preparation of each component for this complex cellular dance. The DNA condenses (it is wrapped around itself tightly to make it a smaller and stronger package) so that it is less susceptible to breakage during movement across the cell. In doing so, most of the DNA becomes transcriptionally inactive. The Golgi bodies and the endoplasmic reticulum begin to break apart into membranous vesicles that can be more easily and evenly distributed across the cell so that both daughter cells receive about the same. The centrosomes (in animal cells) move from their original position near the nucleus toward opposite sides of the cell, to establish the poles of the mitotic spindle.
MPF phosphorylates microtubule motor proteins and microtubule associated proteins (MAPs) to alter the normal microtubule dynamics and allow the massive reorganization into a mitotic spindle to occur. For example, one target of MPF is PRC1, a bundling protein that is inactivated by phosphorylation, thus allowing individual microtubules to move to new locations more easily than a large bundle could. Other effects are inactivation of stabilizing MAPs, which leads to greater lability of microtubules due to increased incidences of catastrophe. The motor protein targets of MPF are in the kinesin family and the phosphorylation is necessary for some of them to bind to the mitotic spindle.
Prometaphase is sometimes considered a separate phase but is also referred to as late prophase, and is primarily defined by the breakup of the nuclear envelope. This process is induced by MPF phosphorylation of the nuclear lamins. Adorned with negative charges from the phosphates, the lamins refuse to associate with one another any longer, leading to the breakdown of the nuclear lamina. As the lamins dissociate, the nuclear envelope remains bound to them, and fragments. This nuclear fragmentation must happen so that the mitotic spindle can reach inside and attach to the chromosomes.
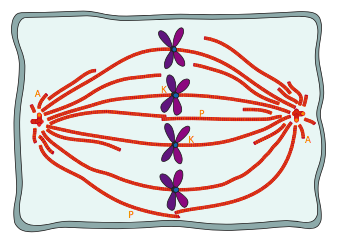
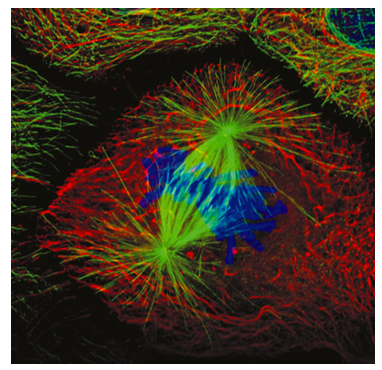
Some of the microtubules of the mitotic spindle attach to the chromosomes via the kinetochore proteins, which link the spindle microtubules to the centromere region of each chromosome. These are known as kinetochore microtubules (Figure \(\PageIndex{8}\)). There are two other types of microtubules in the mitotic spindle (Figure \(\PageIndex{7}\)): the polar microtubules that reach across the cell and interact with one another to help maintain the separation of the centrosomes and defining the overall length of the spindle, and the aster microtubules that are generally short, radiating out from, and stabilizing the centrosome. Remember that the DNA replicated earlier in S phase, and thus sister chromatids are still partially attached. Visually, the centromere region appears narrower or more compressed than the rest of the chromosome, and generally lies near the middle. The centromere contains repeated sequences that are involved in kinetochore binding and assembly.
In primates, the repeating motif is known as alpha satellite DNA, which is made of multiple instances of tandem repeats of a core ~170bp sequence over a centromeric DNA span over a megabase in length. Similar repeats are found in various other vertebrates as well. In other eukaryotes, the size and sequence may vary; for example, much shorter repeats of ~5bp are found in centromeric DNA measuring 200-600kb in Drosophila chromosomes, and S. pombe has centromeric DNA well under 10kb.
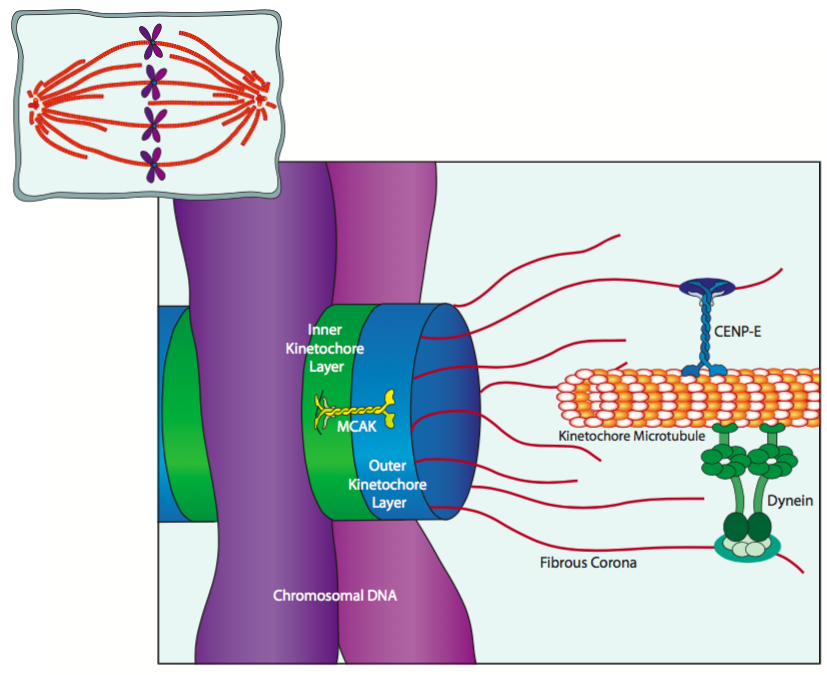
The kinetochores attaching to the centromere DNA are trilaminar protein structures consisting of an inner layer, an outer layer, and a fibrous corona. The kinetochore microtubules of the mitotic spindle are primarily attached to the fibrous corona. As depicted in the Figure, it is attached through CENP-E, a kinesin, and dynein motor proteins that bind along the barrel of the microtubule. In fact, sometimes the first contact between a chromosome (via the kinetochore) and a spindle microtubule is somewhere in the middle of the microtubule, and a combination of microtubule dynamics and motor protein activity move the chromosome to the distal end of the microtubule. This is facilitated by MCAK (mitotic centromere-associated kinesin), which is associated with the kinetochore core proteins and plays a role in depolymerizing microtubules near the (+) end.
The transition from the interphase microtubule cytoskeleton to a mitotic spindle require a number of molecular motors to move the centrosomes, align the microtubules, and expand the spindle. These are depicted in Figure \(\PageIndex{9}\). Initially, as the duplicated centrosomes move away from each other along with some of the cytoskeletal microtubules, the microtubules will interact at various angles. Because the polar microtubules that help to expand or maintain the spindle width must ineract in parallel, cytoplasmic dyneins bind to the eventual polar microtubules and by moving one along the other, bring them into parallel (9a). Once in that position, BimC and other kinesins take over as the primary motors along polar microtubules. They create an outward pushing force by holding onto a microtubule facing one direction, and driving along a parallel MT facing the opposite direction towards the (+) end (9b). Finally, cytosolic dyneins attached to cortical cytoskeleton pull on the astral microtubules, which pulls the spindle ends further from center (9c).
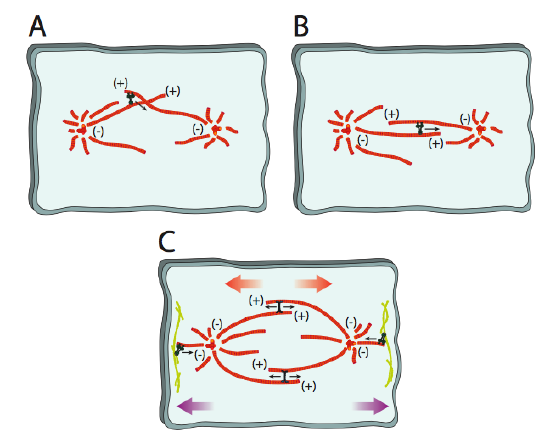
In fact, there appear to be two mechanisms at work: the bub1/bub2 system works in the tension sensing pathway, while another metaphase protein, mad2 appears to be important in suspending mitosis upon disconnection of the kinetochore with the spindle microtubule.
As the nuclear envelope is breaking apart, the mitotic spindle microtubules are undergoing increased dynamic instability, cycling between periods of growth spurts (polymerization) and rapid shortening (catastrophic disassembly), searching for chromosomes to connect to. Once the kinetochore microtubules connect to the chromosomes, the microtubule dynamics shift. The microtubule will primarily undergo shortening if it is beyond the center of the spindle and primarily lengthening if it is short of center. Since eventually each set of sister chromatids is connected to microtubules on both kinetochores, each chromatid is connected to one shortening and one lengthening microtubule. As the chromosomes approach the center of the mitotic spindle, the rate of microtubule shortening/lengthening slows. The sister chromatids are pushed and pulled by the spindle microtubules until they are all lined up along the midline of the mitotic spindle, which in most (but not all) cases is also the midline of the cell. Once they are all lined up, the cell is considered to have reached metaphase. Unlike the other phases, metaphase is a relatively static phase - it is a checkpoint for lining up the chromosomes.
The chromosomes must be properly aligned to ensure that both daughter cells receive the proper complement of chromosomes. How does the cell know when the chromosomes have reached the center of the spindle? An elegantly simple experiment demonstrated that the general mechanism is a tension check - if the two microtubules connecting to the pair of sister chromatids from each side are of the same length, they should be exerting equal tension on the chromosomes. If the microtubule-kinetochore connection is severed at metaphase, the cell will be prevented from progressing (Nicklas, R.B., et al, J. Cell Biol. 130: 929-39, 1995). However, if an equivalent tension is applied by tugging on the chromosome with a glass microneedle, progression of mitosis is restored!
In addition to the tension check, there is another condition that must be met for continuation of mitosis: the MPF must be inactivated. As outlined earlier, MPF in part leads to its own inactivation by activating the anaphase-promoting complex (APC), which polyubiquitinates the cyclin, leading to its destruction and thus MPF-cdk inactivation. APC also tags securin for destruction. Securin is a protein that binds and inhibits the proteolytic enzyme, separase, the activation of which is needed to allow the sister chromatids to separate, which in turn, is necessary for anaphase to proceed.
Barring pathological situations, if and only if the chromosomes all line up at the metaphase plate will the cell proceed to the next stage of mitosis: anaphase. The sister chromatids separate and are pulled toward opposite poles of the mitotic spindle. Somewhat perversely, even as the chromosomes move towards the spindle poles, the poles themselves move outward slightly. Separation of the sister chromatids requires the dissociation of the molecular “glue” holding them together: the cohesin proteins. The cohesins bind to both molecules of DNA and hold them together shortly after replication back in S phase. As anaphase approaches, the enzyme separase is activated, which then cuts the cohesin molecules. Once all of the cohesin molecules are cut, the sister chromatids can finally be separated. The removal of the cohesins proceeds roughly inwards from the distal points of the chromosomes to the centromere, which is generally the last region of attachment.
A cohesin is a multimer of four subunits, Scc1, Scc3, Smc1, and Smc3 in yeast. An additional protein has also been observed in Xenopus. The SCC1 protein is cleaved by separin in yeast, but in metazoans, SCC1 may be removed from chromosomes by another method as well. It is phosphorylated, which decreases its affinity for DNA, and may expose a site for separase-catalyzed hydrolysis.
Separase also promotes anaphase by activating Cdc14, a phosphatase needed to dephosphorylate the cdk substrates that had been phosphorylated by the cyclin-cdk complexes of early mitosis. In addition, Cdc14 is also required for cytokinesis in the yeast S. cerevisiae and nematode C. elegans.
Anaphase can actually be divided into two stages, sometimes referred to as early and late or A and B. At first, the kinetochore microtubules are shortening from both ends, and kinesin-family motors pull the microtubules back toward the spindle poles. As late anaphase starts, polar microtubules elongate, and an additional chromatid-separating force is applied by kinesin-family motor proteins [kinesin-5] that push the polar microtubules against one another to increase the separation between the poles. Dynein-family motors help to direct movement of the poles as well, through their attachment to the aster microtubules and the cortical (peripheral) cytoskeleton.
When both sets of chromosomes arrive at their respective poles, telophase begins. Technically, it was slowly building up since anaphase: when MPF was inactivated by APC, its ability to phosphorylate nuclear lamins was ended. Protein phosphatases in the cell remove the phosphate groups, allowing the lamins to once again interact with one another, and by telophase they are reconstituting the nuclear lamina and the nuclear envelope. Since the lamins and other nuclear membrane proteins also interact with DNA, the nuclear membrane fragments dispersed back in late prophase now coalesce around each set of DNA to form the new nuclear envelopes. The other fragmented membranous organelles (ER, Golgi) also start to re-form. By the end of telophase, the product is a single large cell with two complete nuclei on opposite sides. The next and last step, cytokinesis, splits the cell into two separate and independent daughter cells. In animal cells, cytokinesis is similar to the tightening of a drawstring in the middle of the cell, pulling the “waist” in until all edges meet, and two separate cells result. This contractile ring is composed of actin (structural) and myosin (motive) subunits. These proteins, using ATP for energy, ratchet themselves closer and closer together similar to the actin-myosin “power stroke” described for muscle cell sarcomeres, also primarily made from actin and myosin. This mechanism is universal for animal cells, but the placement of the ring is not always in the center of the cell. The ring often coincides with the center of the cell, but is in fact positioned by the metaphase plate (i.e. the center of mitotic spindle). The most obvious example of a metaphase plate that does not coincide with the center of the cell is found in the formation of egg cells. Because the purpose of an egg cell is to provide all of the material necessary to make a viable new organism upon fertilization (the sperm contributes negligible biomass beyond the genetic material), it divides asymmetrically, with the mitotic spindle located far to one side of the cell (Figure \(\PageIndex{11}\)).
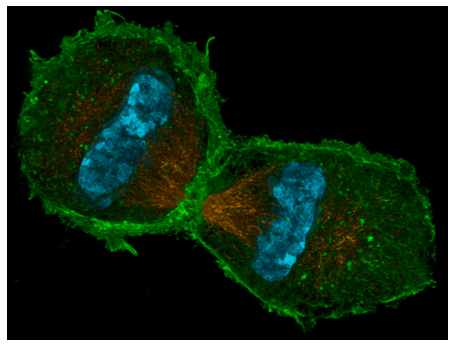
When cytokinesis occurs, one daughter cell, the presumed oocyte, is very large, while the other cell, called a polar body, has minimal cytoplasmic material surrounding the nucleus. The contractile ring works in animal cells because the cell membrane is flexible. In plant cells, the cell membrane is firmly attached to a rigid cell wall, and thus cannot be pulled in. So, the plant cell ingeniously builds a wall down the middle of the cell using specialized vesicles that originate from part of the Golgi, and which contain the materials necessary to form a cell wall. The vesicles travel along the phragmoplast, a structure built from the mitotic spindle microtubules, and as the vesicles line up along the middle of the cell, they begin to fuse to form bigger vesicles and then a large disk-like vesicle, the cell plate. Eventually they reach the cell membrane itself, and fusing with that leads to formation of a new cell wall, and two complete and independent cells.
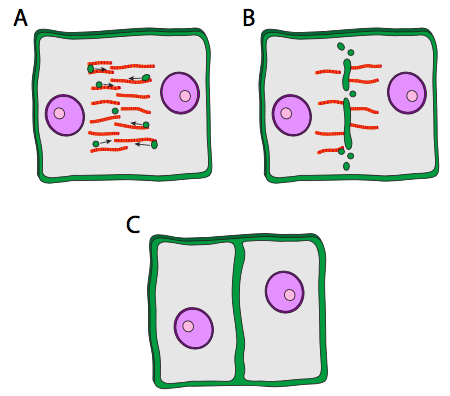
The contents of the vesicles traveling along the phragmoplast are not well described. Callose, a glucose polysaccharide with β1-3 linkages is known to be present in the developing cell plate, but has not been found in the Golgi or vesicles. Interestingly, once the cell plate has fused completely with the existing cell walls, callose gradually disappears. It is thought that the same enzyme system that synthesized callose may switch to synthesizing cellulose as the cell plate matures.