
16.2: Plasma Membrane Structure
- Page ID
- 16514

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)In eukaryotic cells, the plasma membrane surrounds a cytoplasm filled with ribosomes and organelles. Organelles are structures that are themselves encased in membranes. Some organelles (nuclei, mitochondria, chloroplasts) are even surrounded by double membranes. All cellular membranes are composed of two layers of phospholipids embedded with proteins. All are selectively permeable (semi-permeable), allowing only certain substances to cross the membrane. The unique functions of cellular membranes are due to their different phospholipid and protein compositions. Decades of research have revealed these functions (see earlier discussions of mitochondrial and chloroplast function for instance). Here we’ll describe general features of membranes, using the plasma membrane as our example.
A. The Phospholipid Bilayer
Gorter and Grendel predicted the bilayer membrane structure as early as 1925. They knew that red blood cells (erythrocytes) have no nucleus or other organelles, and thus have only a plasma membrane. They also knew that the major chemical component of these membranes were phospholipids. The space-filling molecular model below shows the basic structure of phospholipids, highlighting their hydrophilic (polar) heads and hydrophobic tails.
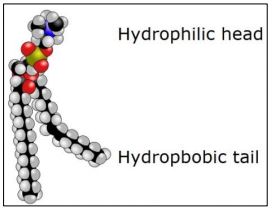
Molecules with hydrophilic and hydrophobic domains are amphipathic molecules. Gorter and Grendel had measured the surface area of red blood cells. They then did a ‘blood count’ and then disrupted a known number of red blood cells. They then measured the amount of phospholipids in the membrane extracts. From this, they calculated that there were enough lipid molecules per cell to wrap around each cell twice. From these observations, they predicted the phospholipid bilayer with fatty acids interacting within the bilayer. Curiously, Gorter and Grendel had made two calculation errors in determining the amount of phospholipid per cells. Nevertheless, their errors compensated each other so that, while not strictly speaking correct, their conclusion remained prophetic! Common membrane phospholipids are shown below.
Amphipathic molecules mixed with water spontaneously aggregate to ‘hide’ their hydrophobic regions from the water. In water, these formed actual structures called liposomes that sediment when centrifuged!
276 Membrane Lipids and the Phospholipid Bilayer
277 Experiments with and Uses of Liposomes
Liposome membrane structure is consistent with the predicted phospholipid bilayer, with the hydrophobic tails interacting with each other and the polar heads facing away from each other, forming a phospholipid bilayer. This led to a picture of membrane architecture based on phospholipid interactions. An iconic illustration of the phospholipid bilayer, with its hydrophobic fatty acid interior and hydrophilic external surfaces is drawn below.
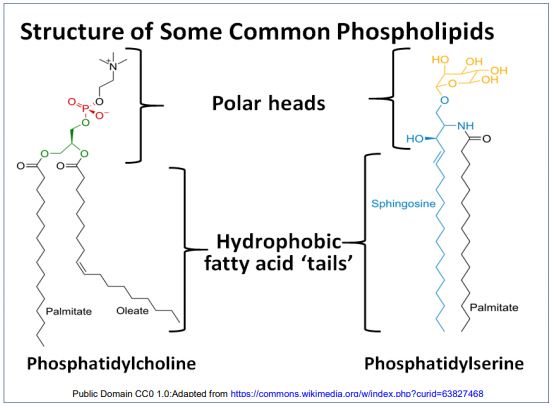
Liposome membrane structure is consistent with the predicted phospholipid bilayer, with the hydrophobic tails interacting with each other and the polar heads facing away from each other, forming a phospholipid bilayer. This led to a picture of membrane architecture based on phospholipid interactions. An iconic illustration of the phospholipid bilayer, with its hydrophobic fatty acid interior and hydrophilic external surfaces is drawn below.

B. Models of Membrane Structure
In 1935, Davson and Danielli suggested that proteins might be bound to the polar heads of the phospholipids in the plasma membrane, creating a protein/lipid/protein sandwich. Decades later, J.D. Robertson observed membranes in the transmission electron microscope at high power, revealing that all cellular membranes had a trilamellar structure. The classic trilamellar appearance of a cellular membrane in the electron microscope is illustrated below
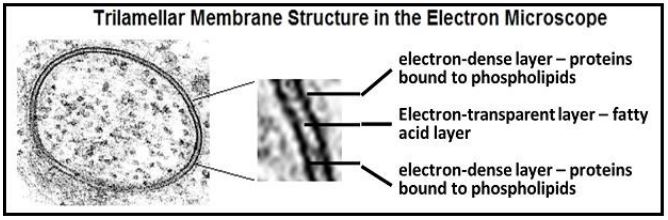
The trilamellar structure is consistent with the protein-coated hydrophilic surfaces of a phospholipid bilayer in Davson and Danielli’s protein-lipid-protein sandwich. Observing that all cellular membranes had this trilamellar structure, Robertson he further proposed his Unit Membrane model: all membranes consist of a clear phospholipid bilayer coated with electron-dense proteins.
The static view of the trilamellar models of membrane structure implied by the Davson-Danielli or Robertson models was replaced in 1972 by Singer and Nicolson’s Fluid Mosaic model (see The fluid mosaic model of membranes. Science 175:720- 731). They suggested that in addition to peripheral proteins that do bind to the surfaces of membranes, many integral membrane proteins actually span the membrane. Integral membrane proteins were imagined as a mosaic of protein ‘tiles’ embedded in a phospholipid medium. But unlike a mosaic of glazed tiles set in a firm, cement-like structure, the protein ‘tiles’ were predicted to be mobile (fluid) in a phospholipid sea. In this model, membrane proteins are anchored in membranes by one or more hydrophobic domains; their hydrophilic domains would face aqueous external and cytosolic environments. Thus, like phospholipids themselves, membrane proteins are amphipathic. We know that cells expose different surface structural (and functional) features to the aqueous environment on opposite sides of a membrane. Therefore, we also say that cellular membranes are asymmetric. A typical model of the plasma membrane of a cell is illustrated below.
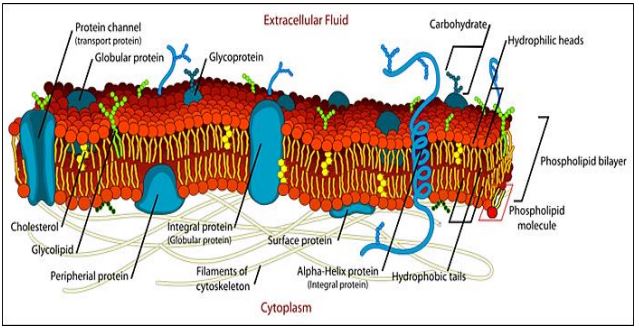
In this model, peripheral proteins have a hydrophobic domain that does not span the membrane, but that anchors it to one side of the membrane. Other peripheral (or socalled “surface”) proteins are bound to the membrane by interactions with the polar phosphate groups of phospholipids, or with the polar domains of integral membrane proteins.
Because of their own aqueous hydrophilic domains, membrane proteins are a natural barrier to the free passage of charged molecules across the membrane. On the other hand, membrane proteins are responsible for the selective permeability of membranes, facilitating the movement of specific molecules in and out of cells. Membrane proteins also account for specific and selective interactions with their extracellular environment. These interactions include the adhesion of cells to each other, their attachment to surfaces, communication between cells (both direct and via hormones and neurons), etc. The ‘sugar coating’ of the extracellular surfaces of plasma membranes comes from oligosaccharides covalently linked to membrane proteins (as glycoproteins) or to phospholipids (as glycolipids). Carbohydrate components of glycosylated membrane proteins inform their function. Thus, glycoproteins enable specific interactions of cells with each other to form tissues. They also allow interaction with extracellular surfaces to which they must adhere. In addition, they figure prominently as part of receptors for many hormones and other chemical communication biomolecules. Protein domains exposed to the cytoplasm, while not glycosylated, often articulate to components of the cytoskeleton, giving cells their shape and allowing cells to change shape when necessary. Many membrane proteins have essential enzymatic features, as we will see. Given the crucial role of proteins and glycoproteins in membrane function, it should come as no surprise that proteins constitute an average of 40-50% of the mass of a membrane. In some cases, proteins are as much as 70% of membrane mass (think cristal membranes in mitochondria!).
278 Properties of Proteins Embedded in a Phospholipid Bilayer
C. Evidence for Membrane Structure
Membrane asymmetry refers to the different membrane features facing opposite sides of the membrane. This was directly demonstrated by the scanning electron microscope technique of freeze-fracture. The technique involves freezing of isolated membranes in water and then chipping the ice. When the ice cracks, the encased membranes split along a line of least resistance… that turns out to be between the hydrophobic fatty acid opposing tails in the interior of the membrane. Scanning electron microscopy then reveals features of the interior and exterior membrane surfaces. Among the prominent features in a scanning micrograph of freeze-fractured plasma membranes are the pits and opposing mounds facing each other on opposite flaps of the membrane, as illustrated below.
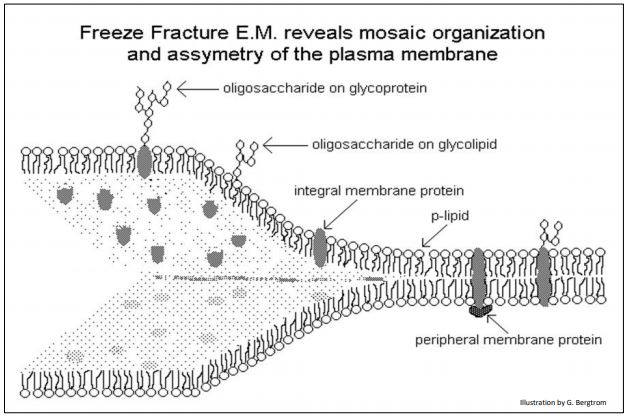
Other features shown here are consistent with phospholipid membrane structure.
280 Freeze Fracture Electron Microscopy of Cell Membranes
Cytochemistry confirmed the asymmetry of the plasma membrane, showing that only the external surfaces of plasma membranes are sugar coated, Check the link below for more detailed descriptions of the experiments.
281 EM Cytochemical Demonstration of Membrane Asymmetry
Finally, the asymmetry of membranes was also demonstrated biochemically. In one experiment, whole cells treated with proteolytic enzymes, followed by extraction of the membranes and then isolation of membrane proteins. In a second experiment, plasma membranes were isolated from untreated cells first, and then treated with the enzymes. In a third experiment, proteins were extracted from plasma membranes isolated from untreated cells. Electrophoretic separation of the three protein extracts by size demonstrated that different components of integral membrane proteins were present in the two digest experiments, confirming the asymmetry of the plasma membrane. Again, for more details, check the link below.
282 Electrophoretic Demonstration of Membrane Asymmetry
The idea that membranes are fluid was also tested. In yet another elegant experiment, antibodies were made to mouse and human cell membrane proteins. Membranes were isolated and injected into a third animal (a rabbit most likely). The rabbit saw the membranes and their associated proteins as foreign and responded by making specific anti-membrane antibody molecules. The antibodies against each membrane source were isolated and separately tagged with different colored fluorescent labels so that they would glow a different color when subjected to ultraviolet light. After mouse and human cells were mixed under conditions that caused them to fuse, making human-mouse hybrid cells. When added to fused cells, the tagged antibodies bound to the cell surface proteins. After a short time, the different fluorescent antibodies were seen to mix under a fluorescence microscope under UV light. The fluorescent tags seemed to moving from their original location in the fused membranes. Clearly, proteins embedded in the membrane are not static, but are able to move laterally in the membrane, in effect floating and diffusing in a “sea of phospholipids”. The mouse antibodies as seen in the hybrid cell right after fusion are cartooned below.
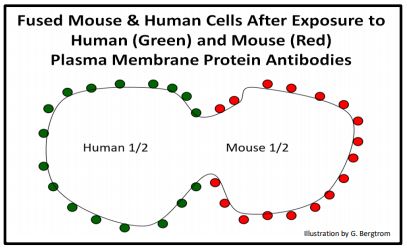
283 Two Demonstrations of Membrane Fluidity: The Fluid Mosaic
D. Membrane Fluidity is Regulated
1. Chemical Factors Affecting Membrane Fluidity
As you might imagine, the fluidity of a membrane depends on its chemical composition and physical conditions surrounding the cell, for example the outside temperature. Factors that affect membrane fluidity are summarized below.

Just as heating a solution causes dissolved molecules and particulates to move faster, phospholipid and protein components of membranes are also more fluid at higher temperatures. If the fatty acids of the phospholipids have more unsaturated (C=C) carbon bonds, these hydrophobic tails will have more kinks, or bends. The kinks tend to push apart the phospholipid tails. With more space between the fatty acid tails, membrane components can move more freely. Thus, more polyunsaturated fatty acids in a membrane make it more fluid. On the other hand, cholesterol molecules tend to fill the space between fatty acids in the hydrophobic interior of the membrane. This reduces the lateral mobility of phospholipid and protein components in the membrane. By reducing fluidity, cholesterol reduces membrane permeability to some ions.
2. Functional Factors Affecting Membrane Fluidity
Evolution has adapted cell membranes to different and changing environments to maintain the fluidity necessary for proper cell function. Poikilothermic, or coldblooded organisms, from prokaryotes to fish and reptiles, do not regulate their body temperatures. Thus, when exposed to lower temperatures, poikilotherms respond by increasing the unsaturated fatty acid content of their cell membranes. At higher temperatures, they increase membrane saturated fatty acid content. Thus, the cell membranes of fish living under the arctic ice maintain fluidity by having high levels of both monounsaturated and polyunsaturated fatty acids. What about fish species that range across warmer and colder environments (or that live in climates with changing seasons). For these fish, membrane composition can change to adjust fluidity to environment.
The warm-blooded (homeothermic) mammals and birds maintain a more or less constant body temperature. As a result, their membrane composition is also relatively constant. But there is a paradox! Their cell membranes are very fluid, with a higher ratio of polyunsaturated fat to monounsaturated fats than say, reptiles. The apparent paradox is resolved however, when we understand that this greater fluidity supports the higher metabolic rate of the warm-blooded species compared to poikilotherms. Just compare the life styles of almost any mammal to a lazy floating alligator, or a snake basking in the shade of a rock!
E. Making and Experimenting with Artificial Membranes
Membrane-like structures can form spontaneously. When phospholipids interact in an aqueous environment, they aggregate to exclude their hydrophobic fatty tails from water, forming micelles. Micelles are spherical phospholipid monolayer vesicles that self-assemble, a natural aggregation of the hydrophobic fatty acid domains of these amphipathic molecules.
A micelle is drawn below.

Micelles can further self-assemble into spherical phospholipid bilayers called liposomes (below).

When formed in the laboratory, these structures behave somewhat like cells, for example, forming a pellet at the bottom of a tube when centrifuged. Liposomes can be custom designed from different kinds of phospholipids and amphipathic proteins that become integral to the liposome membranes. When liposomes can be prepared in the presence of specific proteins or other molecules that can’t cross the membrane. The trapped molecules cannot get out of this synthetic ‘organelle’. Such were the studies that allowed the identification of the mitochondrial respiratory chain complexes. The ability to manipulate liposome content and membrane composition also make them candidates for the drug delivery to specific cells and tissues (google liposome for more information).
F. The Plasma Membrane is Segragated into Regions with Different Properties of Fluidity and Selective Permeability
As we will see shortly, fluidity does not result in an equal diffusion of all membrane components around the cell membrane surface. Instead, extracellular connections between cells as well as intracellular connections of the membrane to differentiated regions of the cytoskeleton, effectively compartmentalize the membrane into subregions. To understand this, imagine a sheet of epithelial like those in the cartoon below.

The sheet of cells exposes one surface with unique functions to the inside of the organ they line. It exposes the opposite surface, one with a quite different function, to the other side of the sheet. The lateral surfaces of the cells are yet another membrane compartment, one that functions to connect and communicate between the cells in the sheet. Components, i.e., membrane proteins illustrated with different symbolic shapes and colors, may remain fluid within a compartment. Of course, this macrodifferentiation of cell membranes to permit cell-cell and cell-environmental interactions makes intuitive sense.
The recent observation that cellular membranes are even more compartmentalized was perhaps less anticipated. In fact, membranes are further divided into microcompartments. Within these compartments, components are fluid but seldom move between compartments. Studies indicate that cytoskeletal elements create and maintain these micro-discontinuities. For example, integral membrane proteins are immobilized in membranes if they are attached to cytoskeletal fibers (e.g., actin) in the cytoplasm. Furthermore, when aggregates of these proteins line up due to similar interactions, they form kind of fence, inhibiting other membrane components from crossing. By analogy, this mechanism of micro-compartmentalization is called the Fences and Pickets model; proteins attached to the cytoskeleton serve as the pickets. The movement across the fences (i.e., from one membrane compartment to another) is infrequent. Extra kinetic energy is presumably needed for a molecule to ‘jump’ a fence between compartments. Hence, this kind of motion, or hop diffusion distinguishes it from the Brownian motion implied by the original fluid mosaic model.
285 Membrane Domains: Regional Differentiation of a Plasma Membrane